- 1Key Laboratory of Forest Ecology and Environment of National Forestry and Grassland Administration, Ecology and Nature Conservation Institute, Chinese Academy of Forestry, Beijing, China
1中国林业科学研究院生态与自然保护研究所, 国家林业和草原局森林生态与环境重点实验室, 中国 北京 - 2Department of Environmental Science and Ecology, School of Environmental Science and Engineering, Shaanxi University of Science and Technology, Xi’an, China
阿拉伯数字陕西科技大学环境科学与工程学院环境科学与生态系,中国 习 - 3Institute of Ecological Conservation and Restoration, Chinese Academy of Forestry, Beijing, China
3中国林业科学研究院生态保护与修复研究所, 中国 北京
Considerable evidences highlight the occurrence of increasing widespread tree mortality as a result of global climate change-associated droughts. However, knowledge about the mechanisms underlying divergent strategies of various tree species to adapt to drought has remained remarkably insufficient. Leaf stomatal regulation and embolism resistance of stem xylem serves as two important strategies for tree species to prevent hydraulic failure and carbon starvation, as comprising interconnected physiological mechanisms underlying drought-induced tree mortality. Hence, the physiological and anatomical determinants of leaf stomatal regulation and stems xylem embolism resistance are evaluated and discussed. In addition, root properties related to drought tolerance are also reviewed. Species with greater investment in leaves and stems tend to maintain stomatal opening and resist stem embolism under drought conditions. The coordination between stomatal regulation and stem embolism resistance are summarized and discussed. Previous studies showed that hydraulic safety margin (HSM, the difference between minimum water potential and that causing xylem dysfunction) is a significant predictor of tree species mortality under drought conditions. Compared with HSM, stomatal safety margin (the difference between water potential at stomatal closure and that causing xylem dysfunction) more directly merge stomatal regulation strategies with xylem hydraulic strategies, illustrating a comprehensive framework to characterize plant response to drought. A combination of plant traits reflecting species’ response and adaptation to drought should be established in the future, and we propose four specific urgent issues as future research priorities.
大量证据强调,由于全球气候变化相关的干旱,树木普遍死亡的发生率不断增加。然而,关于各种树种适应干旱的不同策略背后的机制的了解仍然非常不足。茎木质部的叶片气孔调节和抗栓塞性是树种防止水力衰竭和碳饥饿的两种重要策略,因为它构成了干旱诱导树木死亡的相互关联的生理机制。因此,评价和讨论了叶片气孔调节和茎木质部栓塞抗性的生理和解剖决定因素。此外,还回顾了与耐旱性相关的根特性。在干旱条件下,对叶和茎投入较大的物种往往能保持气孔开放并抵抗茎栓塞。总结并讨论了气孔调节与茎栓塞耐药之间的协调性。先前的研究表明,水力安全边际 (HSM,最小水势与导致木质部功能障碍之间的差异) 是干旱条件下树种死亡率的重要预测指标。与 HSM 相比,气孔安全边际 (气孔关闭时的水势与导致木质部功能障碍的水势之间的差异) 更直接地将气孔调节策略与木质部水力策略相结合,说明了描述植物对干旱响应的综合框架。未来应建立反映物种对干旱的反应和适应的植物性状组合,并提出四个具体的紧迫问题作为未来的研究重点。
Introduction 介绍
Forests, as the dominant biomes of the global land, plays a crucial role in the biogeochemical cycle (Pan et al., 2011). A great amount of evidences highlights increasing forest mortality as a result of global climate change-associated severe drought, and such mortality can lead to large-scale detrimental effect on ecosystem structure and function, causing a conversion from the forests carbon sink into carbon source (Phillips et al., 2009; Hartmann et al., 2018). Global observations of drought-induced forest dieback have stressed a need to understand and predict the vulnerability of trees to more frequent drought in the future.
森林作为全球陆地的主要生物群落,在生物地球化学循环中起着至关重要的作用(Pan et al., 2011)。大量证据表明,由于与全球气候变化相关的严重干旱,森林死亡率增加,这种死亡率可对生态系统结构和功能造成大规模的不利影响,导致森林碳汇转化为碳源(Phillips 等 人,2009 年;Hartmann等 人,2018 年)。对干旱引起的森林枯萎病的全球观测强调了了解和预测树木在未来更频繁的干旱中的脆弱性的必要性。
The mechanisms underlying divergent strategies of tree species to drought have remained remarkably difficult to study, leading to some uncertainty in forecasting the future of forests in the world (Arend et al., 2021). Moreover, differences among tree species in drought adaptability are also integral determinants of distributions and the probability of extinctions in the future (Bonan, 2008). At present, two interconnected physiological reasons underlying drought-induced tree death have evolved, hydraulic failure and carbon starvation (McDowell et al., 2008; McDowell, 2011; Choat et al., 2018; Brodribb et al., 2020).
树种对干旱的不同策略背后的机制仍然非常难以研究,导致预测世界森林的未来存在一些不确定性(Arend 等 人,2021 年)。此外,树种之间在干旱适应性方面的差异也是分布和未来灭绝可能性的不可或缺的决定因素(Bonan,2008 年)。目前,干旱导致树木死亡的两个相互关联的生理原因已经发展出来,即水力衰竭和碳饥饿(McDowell et al., 2008;McDowell, 2011;Choat et al ., 2018;Brodribb等 人,2020 年)。
Based on the cohesion-tension theory, water in plants is transported from the soil through the plant to the crown along a gradient of negative pressure (tension) in the conduits (tracheids or vessels) of the xylem (McDowell et al., 2008; Trugman et al., 2021). Nevertheless, during transpiration, embolism can occur when the pressure in the conduits becomes negative enough to cause air entry through pits into neighboring conduits (Tyree and Zimmermann, 2002). The conduit will become incapable of water transport if embolism occurs (Tyree and Zimmermann, 2002). The hydraulic failure is that drought-induced xylem embolism stops water flow, and subsequently plant tissues desiccate (McDowell et al., 2008). Therefore, tree’ xylem embolism resistance serves as an important strategy to prevent hydraulic failure (Pivovaroff et al., 2016). Another important hydraulic strategy for trees to cope with drought is the closure of stomata. The CO2 used for photosynthesis and the water lost in transpiration share the common pathway—the stomatal pores on leaf surfaces, and reduced transpiration through closuring stomata is inevitably accompanied by the expense of carbon gain. The term “carbon starvation” is that drought induced-stomatal closure (even leaf shedding) causes carbohydrates supply to drop off and the continued carbohydrates metabolism or even the impaired phloem conductance leads to plant tissues starve (McDowell et al., 2008; McDowell, 2011).
基于内聚-张力理论,植物中的水沿着木质部管道(管胞或血管)中的负压(张力)梯度从土壤通过植物输送到树冠(McDowell et al., 2008;Trugman等 人,2021 年)。然而,在蒸腾过程中,当导管中的压力变得足够负,导致空气通过凹坑进入相邻的导管时,就会发生栓塞(Tyree 和 Zimmermann,2002 年)。如果发生栓塞,导管将无法进行水运输(Tyree 和 Zimmermann,2002 年)。水力故障是干旱引起的木质部栓塞停止水流,随后植物组织干燥(McDowell et al., 2008)。因此,树的木质部栓塞阻力是防止水力衰竭的重要策略(Pivovaroff et al., 2016)。树木应对干旱的另一个重要水力策略是关闭气孔。用于光合作用的 CO2 和蒸腾作用中损失的水分具有共同的途径——叶片表面的气孔孔,通过闭气孔减少的蒸腾作用不可避免地伴随着碳增加的代价。术语“碳饥饿”是指干旱诱导的气孔关闭(甚至落叶)导致碳水化合物供应下降,持续的碳水化合物代谢甚至韧皮部电导受损导致植物组织饥饿(McDowell et al., 2008;McDowell,2011 年)。
Therefore, xylem embolism resistance and stomatal regulation are closely related to hydraulic failure and carbon starvation. In other words, the risk of tree mortality is generally determined by the lethal thresholds of carbon depletion and/or hydraulic failure to which plants are exposed during drought. The lethal thresholds are mainly the thresholds of embolism resistance and stomatal regulation (McDowell et al., 2008; Choat et al., 2018; Blackman et al., 2019; Brodribb et al., 2020). In fact, previous studies have indeed shown that drought-induced mortality of a given species can be predicted by hydraulic thresholds (xylem pressure at which 50% loss of hydraulic conductance, P50, a hydraulic trait widely used to estimate hydraulic safety) and declining carbon availability (Choat et al., 2012; Anderegg et al., 2016; Anderegg et al., 2019 Chen et al., 2019a).
因此,木质部栓塞抵抗力和气孔调节与水力衰竭和缺碳密切相关。换句话说,树木死亡的风险通常由植物在干旱期间面临的碳消耗和/或水力衰竭的致命阈值决定。致死阈值主要是栓塞抵抗和气孔调节的阈值(McDowell et al., 2008;Choat et al ., 2018;Blackman et al., 2019;Brodribb等 人,2020 年)。事实上,以前的研究确实表明,干旱引起的给定物种的死亡率可以通过水力阈值(水力传导损失 50% 的木质部压力,P50,一种广泛用于估计水力安全性的水力特性)和碳可用性的下降来预测(Choat 等 人,2012 年;Anderegg et al., 2016;Anderegg et al., 2019Chen et al., 2019a)。
The water is limited in the soil, and the ability of terrestrial plants to resist embolism and regulate stoma is limited. A conifer species from arid regions of Western Australia, Callitris tuberculata, is the most embolism resistant tree species in the world to date (P50 = -18.8 MPa) (Larter et al., 2015). In addition, stomatal regulation is also a requisite condition for land plants evolved to adapt to the terrestrial environment and it is impossible that endless water from the soil to be transpired into the atmosphere. Therefore, tree species with different adaptation strategies are screened and survived in different environments, which also achieves colorful biodiversity in terrestrial environments with various moisture conditions.
土壤中的水分有限,陆生植物抵抗栓塞和调节造口的能力是有限的。来自西澳大利亚干旱地区的针叶树种 Callitris tuberculata 是迄今为止世界上最抗栓塞的树种 (P50 = -18.8 MPa) (Larter et al., 2015)。此外,气孔调节也是陆地植物进化以适应陆地环境的必要条件,土壤中无尽的水不可能蒸腾到大气中。因此,筛选具有不同适应策略的树种,并在不同的环境中生存,这也在各种水分条件下的陆地环境中实现了丰富多彩的生物多样性。
In this review, we evaluate and discuss the physiological and anatomical determinant of xylem embolism resistance and leaf stomatal regulation. The coordination and trade-offs of the two drought adaptation strategies are also summarized. Root, as important organ for absorbing water and nutrients, and its coordination with leaf and stem are also reviewed. In addition, research prospects of trees’ drought adaptation strategies in the future are proposed.
在这篇综述中,我们评估和讨论了木质部栓塞耐药和叶片气孔调节的生理和解剖决定因素。本文还总结了两种干旱适应策略的协调和权衡。还综述了根作为吸收水分和养分的重要器官,及其与叶和茎的协调性。此外,还对未来树木的干旱适应策略的研究前景提出了展望。
Stomatal regulation 气孔调节
Based on fossil record, stomata appear approximately 400 million years ago (Edwards et al., 1998; Brodribb and Holbrook, 2003). The first published stomatal measurement was about stomatal density (Sack and Buckley, 2016), and the recognition that stomata open and close by changes in guard cell turgor was in 1938 (Heath, 1938). Although the total area of stomatal pores occupies only a small fraction of the leaf surface, typically less than 3%, 98% of H2O and CO2 passes through these pores (Lawson and Blatt, 2014). The evolution of stoma is believed to be caused by the selective pressure of optimizing the ratio of CO2 uptake to water lost during gas exchange (Raven, 2002; Brodribb and Holbrook, 2003). When plants experience drought, stomatal closure will be the first response of plant to reduce transpiration (Bartlett et al., 2016).
根据化石记录,气孔出现在大约 4 亿年前(Edwards 等 人,1998 年;Brodribb 和 Holbrook,2003 年)。首次发表的气孔测量是关于气孔密度的 (Sack and Buckley, 2016),并且认识到气孔通过保卫细胞膨胀的变化打开和关闭是在 1938 年(Heath, 1938)。虽然气孔孔的总面积只占叶表面的一小部分,通常不到 3%,但 98% 的 H2O 和 CO2 通过这些孔(Lawson 和 Blatt,2014)。造口的演变被认为是由优化 CO2 吸收与气体交换过程中损失的水的比率的选择压力引起的(Raven,2002 年;Brodribb 和 Holbrook,2003 年)。当植物经历干旱时,气孔关闭将是植物减少蒸腾作用的第一反应(Bartlett et al., 2016)。
Stomal opening or closing is controlled by inflating or deflating stomatal guard cells driven by influx or efflux of water, whose water potential is associated with chemical osmolytes or signals (such as pH, ABA, H+, K+, Cl2 and ethephon, etc.) and leaf water potential (Brodribb et al., 2014; Medeiros et al., 2015; Papanatsiou et al., 2019). Therefore, water potential plays a key role in determining stomatal movements. In addition, guard cell size (and geometry) also tends to affect the speed of stomatal movement, and larger stomata often exhibit slower responses (Lawson and Blatt, 2014). In this review, the summarization and discussion will focus on the effect of water status on stomatal opening and closure.
气孔的打开或关闭是通过水的流入或流出驱动的气孔守卫细胞的充气或放气来控制的,其水势与化学渗透物或信号(如 pH、ABA、H+、K+、Cl2 和乙烯利等)和叶片水势有关(Brodribb等 ,2014 年;Medeiros等 人,2015 年;Papanatsiou等 人,2019 年)。因此,水势在决定气孔运动中起着关键作用。此外,保卫细胞的大小(和几何形状)也往往影响气孔运动的速度,并且较大的气孔通常表现出较慢的反应(Lawson 和 Blatt,2014)。在这篇综述中,总结和讨论将集中在水状态对气孔打开和关闭的影响。
The concept of isohydry and anisohydry
isohydry 和 anisohydry 的概念
Significant variation in sensitivity and responsiveness of the opening and closing of stomata to external environmental and internal signaling cues to exist among species (Lawson and Blatt, 2014). Further, the species-specific differences in the relationship between stomatal behavior and dynamic variation in water conditions are important for understanding the species-specific differences in the performance of response and adaptation to the environment (Meinzer et al., 2016). Frequently, across species, stomatal regulation is classified along a continuum from isohydry to anisohydry (Brodribb and McAdam, 2013; Brodribb et al., 2014; Skelton et al., 2015; Meinzer et al., 2016; Chen et al., 2021b; Hartmann et al., 2021). This classification existed before the water potential concept (Slatyer and Taylor, 1960; Scholander et al., 1965) and initially focused more on the regulation of transpiration than on the maintaining stable leaf water potential (Ψleaf) per se (Martínez-Vilalta and Garcia-Forner, 2017). Now the definition of iso/anisohydric is usually based on Ψleaf, and anisohydric species is generally attributed to less stomatal sensitivity, allowing large variation in Ψleaf; isohydric species typically exhibit strong stomatal regulation, leading to relatively stable Ψleaf (Tardieu and Simonneau, 1998; Klein, 2014; Meinzer et al., 2017).
气孔打开和关闭对物种之间存在的外部环境和内部信号线索的敏感性和响应性存在显着差异(Lawson 和 Blatt,2014 年)。此外,气孔行为与水条件动态变化之间关系的物种特异性差异对于理解对环境的反应和适应性能的物种特异性差异非常重要(Meinzer et al., 2016)。通常,在物种之间,气孔调节按照从等水态到非水态的连续体进行分类(Brodribb 和 McAdam,2013 年;Brodribb等 人,2014 年;Skelton等 人,2015 年;Meinzer et al., 2016;Chen等 人,2021b;Hartmann et al., 2021)。这种分类在水势概念之前就已经存在(Slatyer 和 Taylor,1960 年;Scholander 等 人,1965 年)最初更多地关注蒸腾作用的调节,而不是维持稳定的叶片水势(Ψ叶)本身(Martínez-Vilalta 和 Garcia-Forner,2017 年)。现在等水态/各水态的定义通常基于 Ψ叶,而非水态物质通常归因于气孔敏感性较低,允许 Ψ叶发生较大变化;等水物质通常表现出强烈的气孔调节,导致相对稳定的 Ψ叶(Tardieu 和 Simonneau,1998 年;Klein,2014 年;Meinzer et al., 2017)。
The quantification of iso/anisohydric behavior
iso/anisohydric 行为的量化
Three methods are generally adopted for the quantification of iso/anisohydric behavior: (1) the relationship between stomatal conductance (Gs) and Ψleaf; (2) the relationship between predawn water potential (Ψpd) and midday water potential (Ψmd); (3) the “hydroscape” method.
通常采用三种方法来量化等/各水力学行为:(1) 气孔导度 (Gs) 和 Ψ叶之间的关系;(2) 黎明前水势 (Ψpd) 与正午水势 (Ψmd) 之间的关系;(3) “水景”方法。
The quantification of iso/anisohydric behavior by definition refers to ascertain the sustainability of Gs in the decrease of Ψleaf during drought, so the water potential at stomatal closure (Ψclose) obtained by the Gs-Ψleaf curve is a suitably quantitative indicator of iso/anisohydric behavior (Brodribb and Holbrook, 2003; Skelton et al., 2015; Chen et al., 2021b; Bartlett and Sinclair, 2021a). This Gs-Ψleaf curve method need to measure a wide enough range of soil moisture or water potential and relevant Gs to construct intact curve for obtaining accurate Ψclose. In addition, Gs is also influenced by CO2 concentration, illumination, air temperature, etc., so these interference factors on Gs must be excluded as much as possible during the measurement. By analyzing Ψclose, many studies found that species-specific Ψclose formed a continuum, not simply dichotomy between isohydric and anisohydric (Klein, 2014; Martin-StPaul et al., 2017; Pivovaroff et al., 2018; Chen et al., 2019a; Henry et al., 2019). In a study of 20 co-occurring temperate broadleaf tree species, Ψclose was found to be in the range from -0.655 MPa to -5.54 MPa (Chen et al., 2019a). In a meta-analysis study gathered data from more than 100 species in different biomes, Ψclose varied from -1 to -4.3 MPa (Martin-StPaul et al., 2017). Noticeably, in a study conducted in southern California with a Mediterranean-type climate, Ψclose was found to be about -10 MPa in a chaparral species (Pivovaroff et al., 2018).
根据定义,等/各水力学行为的量化是指确定干旱期间 Ψ叶片减少中 Gs 的可持续性,因此通过 Gs-Ψ 叶片曲线获得的气孔关闭(Ψ关闭)的水势是等/各水力学行为的适当定量指标(Brodribb 和 Holbrook,2003 年;Skelton等 人,2015 年;Chen等 人,2021b;Bartlett 和 Sinclair,2021a)。这种 Gs-Ψ 叶曲线方法需要测量足够宽的土壤水分或水势范围以及相关的 Gs,以构建完整的曲线以获得准确的 Ψ接近。此外,Gs 还受到 CO2 浓度、光照、空气温度等的影响,因此在测量过程中必须尽可能排除这些对 Gs 的干扰因素。通过分析 Ψclose,许多研究发现物种特异性 Ψclose 形成了一个连续体,而不仅仅是等水和各水之间的二分法(Klein,2014 年;Martin-StPaul et al., 2017;Pivovaroff等 人,2018 年;Chen等 人,2019a;Henry等 人,2019 年)。在对 20 种共存的温带阔叶树种的研究中,发现 Ψclose 在 -0.655 MPa 至 -5.54 MPa 的范围内(Chen et al., 2019a)。在一项荟萃分析研究中,收集了来自不同生物群落中 100 多个物种的数据,Ψclose 从 -1 到 -4.3 MPa 不等(Martin-StPaul et al., 2017)。 值得注意的是,在地中海型气候的南加利福尼亚进行的一项研究中,发现 Chaparral 物种的 Ψclose 约为 -10 MPa(Pivovaroff 等 人,2018 年)。
More isohydric species show less variability in Ψmd, but Ψmd of anisohydric tree species present oppositely (Martínez-Vilalta and Garcia-Forner, 2017). Based on this definition, Martínez-Vilalta et al. (2014) classified the stomatal regulation behaviors of plants by the decline range of Ψmd with the decrease of Ψpd, i.e., the slope (σ) of the model. However, σ vary with soil water potential, which highlights that confirming iso/anisohydric regulation on the basis of differences between predawn and midday water potential is problematic (Hochberg et al., 2018). Furthermore, changes in Ψmd and Ψpd are also affected by hydraulic conductance and Gs, and the reductions in both have the opposite effects on Ψleaf (Martínez-Vilalta and Garcia-Forner, 2017). Thus, considering the large effect of the environment, for proper the degree of iso-anisohydry of the species’ genotypic effect, basic physiological characteristics (such as Ψclose or its proxy, turgor loss point) should be adopted rather than the degree of relationship between Ψpd and Ψmd (Hochberg et al., 2018).
较多的等水物质在 Ψmd 中的变异性较小,但各水性树种的 Ψmd 则相反(Martínez-Vilalta 和 Garcia-Forner,2017 年)。基于这一定义,Martínez-Vilalta 等 人 (2014) 根据 Ψmd 的下降范围和 Ψpd 的降低对植物的气孔调节行为进行分类,即模型的斜率 (σ)。然而,σ随土壤水势而变化,这凸显出根据黎明前和正午水势之间的差异来确认等水/等水调节是有问题的(Hochberg et al., 2018)。此外,Ψmd 和 Ψpd 的变化也受水力电导率和 Gs 的影响,两者的减少对 Ψ叶片有相反的影响(Martínez-Vilalta 和 Garcia-Forner,2017)。因此,考虑到环境的巨大影响,为了适当地确定物种基因型效应的等等水陆程度,应采用基本生理特性(例如 Ψclose 或其代理、膨胀损失点),而不是 Ψpd 和 Ψmd 之间的关系程度(Hochberg et al., 2018)。
Meinzer et al. (2016) coined a new metric, the “hydroscape”, to define tree species’ isohydricity. The hydroscape is a two-dimensional metric, defined as the area enclosed by the Ψpd and Ψmd regression lines, and species with larger hydroscapes are more anisohydric. The hydroscape integrates more comprehensive information and more accurately quantifies the iso/anisohydric degree than simple metrics such as σ (Meinzer et al., 2016). Furthermore, the strong correlation between hydroscape and hydraulic traits also suggests that the hydroscape may provide more information about overall drought tolerance than metrics involving the rate of change in Ψmd as Ψpd declines (Meinzer et al., 2016; Li et al., 2019).
Meinzer 等 人(2016 年)创造了一个新的指标,即“水景”,来定义树种的等水性。水景是一个二维度量,定义为 Ψpd 和 Ψmd 回归线所包围的区域,水景较大的物种更不合水。与σ等简单指标相比,水文景观整合了更全面的信息,并更准确地量化了等离子/非水化度(Meinzer et al., 2016)。此外,水景和水力特征之间的强相关性也表明,水景可能比涉及 Ψmd 随 Ψpd 下降而变化率的指标提供更多关于整体耐旱性的信息(Meinzer et al., 2016;Li et al., 2019)。
In addition to the three mentioned main methods, turgor loss point (TLP) was also regarded as a potential proxy for species stomatal regulation strategy (Hochberg et al., 2018), which is directly and closely linked with stomatal regulation (Brodribb and Holbrook, 2003; Chen et al., 2021b; Figure 1).
除了上述三种主要方法外,膨胀损失点 (TLP) 也被视为物种气孔调节策略的潜在代理(Hochberg et al., 2018),它与气孔调节直接而紧密相关(Brodribb 和 Holbrook,2003 年;Chen等 人,2021b;图 1)。
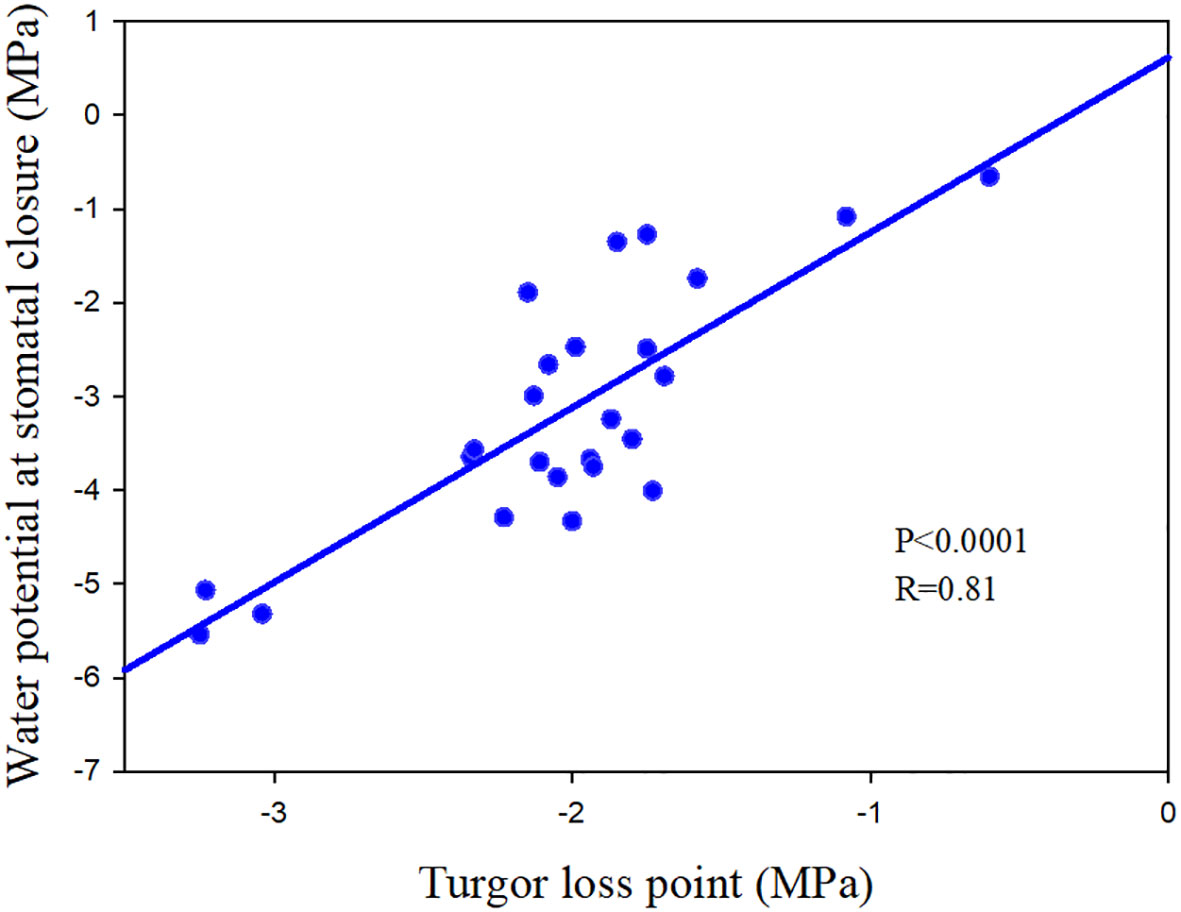
Figure 1 The relationship between turgor loss point and the water potential at stomatal closure of 25 angiosperm speices. The data comes from Chen et al. (2021b) and Li et al. (2015).
图 125 个被子植物特质气孔闭合时膨胀损失点与水势之间的关系。数据来自 Chen 等 人 (2021b) 和 Li 等 人 (2015)。
The determinants of iso/anisohydric behavior
iso/anisohydric 行为的决定因素
Regardless of how iso/anisohydric behavior is defined, it has been shown to be associated with a variety of stem and leaf traits (Fu et al., 2019; Chen et al., 2021b). The determinants or relevant traits of stomatal behavior are mainly phytohormone ABA, hydraulic traits and structural investment as discussed below.
无论如何定义等水/各水行为,它已被证明与各种茎和叶性状相关(Fu et al., 2019;Chen等 人,2021b)。气孔行为的决定因素或相关性状主要是植物激素 ABA、水力性状和结构投资,如下所述。
ABA 阿坝
ABA can cause a reduction in guard cell turgor, subsequently closing stomata (Brodribb and McAdam, 2013). However, the earliest stomatal response was an passive-hydraulic regulation without the requirement of ABA. This can be found in the extant species of basal vascular plants, including the lycophytes, ferns, and gymnosperms in evolutionary terms (McAdam and Brodribb, 2015). The passive-hydraulic regulation in these species show that stomata are like valves closing or opening in response to, respectively, a decrease or increase in turgor (McAdam and Brodribb, 2015).
ABA 可导致保卫细胞弹性减少,随后关闭气孔(Brodribb 和 McAdam,2013 年)。然而,最早的气孔反应是无 ABA 要求的被动液压调节。这可以在现存的基底维管植物物种中找到,包括进化方面的石松植物、蕨类植物和裸子植物(McAdam 和 Brodribb,2015)。这些物种的被动液压调节表明,气孔就像瓣膜一样,分别响应膨胀的减少或增加而关闭或打开(McAdam 和 Brodribb,2015)。
The traditional studies suggest that ABA is a root-related hormone which is transported to the foliage through the stem xylem (Davies and Zhang, 1991), but short-term responses of ABA cannot be made clear by xylem transport (Lee et al., 2006). Now mounting molecular evidence illustrates the capacity of foliage and even guard cells to generate ABA (Bauer et al., 2013), and the ABA concentration may increase after short-term exposure of foliage to drought (Waadt et al., 2014; McAdam and Brodribb, 2015). In addition, the vascular tissues, including the phloem cells, can also synthesize ABA and transport ABA to the guard cells (Okamoto et al., 2009; Kuromori et al., 2014).
传统研究表明,ABA 是一种与根相关的激素,通过茎木质部运输到叶子上(Davies 和 Zhang,1991),但木质部运输无法明确 ABA 的短期反应(Lee et al., 2006)。现在越来越多的分子证据表明,叶子甚至保卫细胞产生 ABA 的能力(Bauer等 人,2013 年),并且叶子短期暴露于干旱后 ABA 浓度可能会增加(Waadt等 人,2014 年;McAdam 和 Brodribb,2015 年)。此外,包括韧皮部细胞在内的维管组织也可以合成 ABA 并将 ABA 转运到保卫细胞(Okamoto et al., 2009;Kuromori et al., 2014)。
Tardieu and Simonneau (1998) proposed that qualitative differences in ABA may be the source of the differences between anisohydric and isohydric plants. Among conifers, contrasting iso/anisohydric behaviors were found to be related to divergent trends in xylem embolism resistance and foliar ABA dynamics during drought, with a functional phylogeny type (Brodribb et al., 2014). The rise in ABA in the xylem or leaf can activate anion channels, causing a reduction of guard cell turgor and subsequent stomatal closure. This mode fits with some species, whose stomata were sensitive to ABA as drought stress intensified, but not fit with other species (Brodribb and McAdam, 2013).
Tardieu 和 Simonneau (1998) 提出,ABA 的质的差异可能是各水合植物和等水合植物之间差异的根源。在针叶树中,发现对比鲜明的等/等水行为与干旱期间木质部栓塞抗性和叶面 ABA 动力学的不同趋势有关,具有功能性系统发育类型(Brodribb et al., 2014)。木质部或叶片中 ABA 的增加可以激活阴离子通道,导致保卫细胞膨胀减少和随后的气孔关闭。这种模式适用于一些物种,随着干旱胁迫的加剧,它们的气孔对 ABA 敏感,但不适合其他物种(Brodribb 和 McAdam,2013)。
The physiological evidences from 42 species of conifers showed that the evolutionary mechanism of stomatal regulation of gymnosperm species follows two distinct pathways (Brodribb et al., 2014). In gymnosperm species with isohydric behavior, foliar ABA levels rapidly rise during drought stress exposure, and high levels of foliar ABA induce complete stomatal closure at relatively high water potential (ABA-rising type) (Brodribb et al., 2014; Kollist et al., 2014). However, in some gymnosperm species with anisohydric behavior, stomatal initial closure is induced by increased ABA and then by a transition to water potential during drought, and the stomata could only be completely closed when the negative water potential pull the guard cell turgor low enough (Brodribb and McAdam, 2013; Brodribb et al., 2014). The similar results have been also obtained in angiosperms, and thus the relationship between ABA dynamics and iso/anisohydric behaviors may be commonplace in woody plants (Brodribb and McAdam, 2013; Nolan et al., 2017). Given that ABA is likely to be an important determinant of stomatal regulation type, the iso/anisohydric behaviors may be regulated by relatively few genes, namely those regulating ABA catabolism or synthesis (Brodribb and McAdam, 2013; Brodribb and McAdam, 2015).
来自 42 种针叶树的生理证据表明,裸子植物物种气孔调节的进化机制遵循两种不同的途径(Brodribb et al., 2014)。在具有等水行为的裸子植物物种中,叶面 ABA 水平在干旱胁迫暴露期间迅速上升,高水平的叶面 ABA 在相对较高的水势下诱导气孔完全关闭(ABA 上升型)(Brodribb et al., 2014;Kollist等 人,2014 年)。然而,在一些具有不水行为的裸子植物物种中,气孔初始关闭是由 ABA 增加诱导的,然后是由干旱期间向水势的转变诱导的,只有当负水势将保卫细胞膨胀拉得足够低时,气孔才能完全关闭(Brodribb 和 McAdam,2013 年;Brodribb et al., 2014)。在被子植物中也获得了类似的结果,因此 ABA 动力学与等/各水力学行为之间的关系在木本植物中可能很常见(Brodribb 和 McAdam,2013 年;Nolan et al., 2017)。鉴于 ABA 可能是气孔调节类型的重要决定因素,iso/iso-hydric 行为可能受相对较少的基因调节,即调节 ABA 分解代谢或合成的基因(Brodribb 和 McAdam,2013 年;Brodribb 和 McAdam,2015 年)。
Hydraulic traits 水力特性
Around the plant water use strategy of maximizing carbon gain and minimizing water loss, there are a set of hydraulic traits coordinated with iso/anisohydric behavior, which is not only the consequence of hydraulic regulation but also related to costs and gains.
围绕最大化碳增益和最小化水分损失的植物用水策略,有一组与 iso/各水力学行为相协调的水力特性,这不仅是水力调节的结果,还与成本和收益有关。
Stomatal behavior directly affects plant water potential, and in previous research minimum midday water potential was strongly correlated with Ψclose, suggesting that Ψclose crucially affects water stress the leaves experience (Bartlett et al., 2016). TLP is an important trait reflecting drought stress tolerance of tree species (Zhu et al., 2018), and stomatal behavior is largely determined by leaf turgor (Figure 1). Lenz et al. (2006) found that TLP was more negative in species inhabited in lower rainfall sites and whose diurnal fluctuations in water potential were larger. Other leaf pressure–volume (P-V) traits, such as relative water content at TLP (RWCtlp) and osmotic potential at full turgor (Ψ100), were found to be strongly related to TLP and consequentially Ψclose. Meinzer et al. (2016) demonstrated that TLP and Ψ100 could serve as robust proxies for a species’ location along the iso/anisohydry continuum within a diverse group of woody species studied under similar conditions. Estimates of TLP and Ψ100 derived from osmometer measurement method could further rapidly streamline rankings of species’ iso/anisohydric stringency (Bartlett et al., 2012b).
气孔行为直接影响植物的水势,在以前的研究中,最小正午水势与 Ψclose 密切相关,这表明 Ψclose 对叶片经历的水分胁迫至关重要(Bartlett et al., 2016)。TLP 是反映树种耐旱性的重要性状(Zhu et al., 2018),气孔行为主要取决于叶片的膨胀度(图 1)。Lenz 等 人(2006 年)发现,生活在降雨量较低地点的物种的 TLP 更为负,其水势的昼夜波动较大。其他叶片压力-体积 (P-V) 性状,例如 TLP 的相对含水量 (RWCtlp) 和完全膨胀时的渗透电位 (Ψ100),被发现与 TLP 密切相关,因此 Ψ接近。Meinzer 等 人(2016 年)证明,TLP 和 Ψ100 可以作为在类似条件下研究的不同木本物种群中物种沿等水态/各水合体位置的可靠代理。从渗透压计测量方法得出的 TLP 和 Ψ100 的估计值可以进一步快速简化物种的 iso/isosohydric 严格度的排名(Bartlett et al., 2012b)。
The TLP is controlled by the accumulation of actively osmotic solutes (Kozlowski and Pallardy, 2002). A meta-analysis study found that Ψ100 was the main determinant of TLP (Bartlett et al., 2012a). In addition to Ψ100, the TLP is always related to the leaf modulus of elasticity (ϵ) (Meinzer et al., 2014). More negative Ψ100 and greater ϵ can contribute to the turgor maintenance as water content and water potential declines, which can maintain leaf physiological activity by maintaining protoplast volume, thus extending stomatal opening and photosynthesis with drying conditions (Meinzer et al., 2014; Johnson et al., 2018). Because stomata generally close around the TLP, stomata rarely closing at very negative water potential may reflect the finiteness of osmotic adjustment and the inability of foliage to maintain turgor at very negative Ψleaf (Bartlett et al., 2012a; Henry et al., 2019). Therefore, the position of species along the isohydric to anisohydric continuum would be scaled with species-specific variation in foliage osmotic properties (Meinzer et al., 2016; Hartmann et al., 2021).
TLP 受主动渗透溶质积累的控制(Kozlowski 和 Pallardy,2002 年)。一项荟萃分析研究发现,Ψ100 是 TLP 的主要决定因素(Bartlett et al., 2012a)。除了 Ψ100 之外,TLP 总是与叶片弹性模量 (ε) 有关(Meinzer et al., 2014)。随着含水量和水势的下降,更多的负 Ψ100 和更大的 ε 有助于维持膨胀,这可以通过维持原生质体体积来维持叶片生理活性,从而在干燥条件下延长气孔开放和光合作用(Meinzer et al., 2014;Johnson et al., 2018)。因为气孔通常在 TLP 周围闭合,所以气孔很少在非常负的水势时闭合,这可能反映了渗透调节的有限性和叶子在非常负的 Ψ叶下无法保持膨胀(Bartlett 等 人,2012a;Henry等 人,2019 年)。因此,物种沿等水到各水连续体的位置将根据叶子渗透特性的物种特异性变化进行缩放(Meinzer et al., 2016;Hartmann et al., 2021)。
The contribution of highly negative osmotic potentials to maintaining turgor is equivalent to more investment in high concentrations of compatible solutes (Johnson et al., 2018; Hartmann et al., 2021). The leaves of isohydric species would not undergo osmoregulation to maintain turgor during drought, which would decrease energy costs related to solute accumulation (Meinzer et al., 2014; Hartmann et al., 2021). However, excessive solute accumulation may result in the deleterious effects on leaf protein activity, which lead to a physiological limit to stomata opening, so osmotic potential is predicted to below the values of Ψclose (Martin-StPaul et al., 2017).
高度负渗透电位对维持膨胀的贡献相当于对高浓度相容溶质的更多投资(Johnson等 人,2018 年;Hartmann et al., 2021)。等水物质的叶子在干旱期间不会进行渗透调节以保持膨胀,这将降低与溶质积累相关的能源成本(Meinzer et al., 2014;Hartmann et al., 2021)。然而,过量的溶质积累可能会对叶片蛋白活性产生有害影响,从而导致气孔张开的生理限制,因此预计渗透电位将低于 Ψclose 的值(Martin-StPaul等 人,2017 年)。
Stomatal conductance is directly linked to leaf hydraulic conductance (Kleaf), and stomatal closure have been shown to be related to a loss of Kleaf, aiming to prevent leaf desiccation (Brodribb and Holbrook, 2003; Woodruff et al., 2015). The coordination between Ψleaf50 (Ψleaf at 50% loss of conductivity) and TLP suggests that decline in Kleaf under drought should partially be a result of mesophyll cells losing turgor and shrinking (Scoffoni et al., 2014). Species with more negative TLP undergo less cell shrinkage and have slower declines in Kleaf during drought (Bartlett et al., 2016). Therefore, the Kleaf of more anisohydric species may be less vulnerable to drought (Nardini and Luglio, 2014), and more isohydric species were found to have lower hydraulic capacitance of leaves but higher Kleaf (Fu et al., 2019). Water storage through capacitance is also a factor in Ψclose (Hammond and Adams, 2019). Nevertheless, there is no consistent conclusion on the relationship between Ψclose and maximum Gs (Meinzer et al., 2017; Henry et al., 2019). Foliar water uptake was found to be an important water acquisition mechanism that can mitigate water deficits in some tree species, but the role of foliar water uptake on stomatal regulation remains unknown (Eller et al., 2013).
气孔导度与叶片水力导度(K叶)直接相关,气孔关闭已被证明与 K叶的损失有关,旨在防止叶片干燥(Brodribb 和 Holbrook,2003 年;Woodruff et al., 2015)。Ψ叶 50 (Ψ叶的电导率损失 50%)和 TLP 之间的协调表明,干旱下 K叶的下降应该部分是由于叶肉细胞失去膨胀和收缩的结果(Scoffoni等 人,2014 年)。在干旱期间,具有更多负 TLP 的物种经历较少的细胞收缩并且 K叶下降较慢(Bartlett等 人,2016 年)。因此,更多非水性物种的 K叶可能不太容易受到干旱的影响(Nardini 和 Luglio,2014),并且发现更多的等氢物种具有较低的叶片水力容量但较高的 K叶(Fu et al., 2019)。通过电容储水也是 Ψclose 的一个因素(Hammond 和 Adams,2019 年)。然而,关于 Ψ接近和最大 Gs 之间的关系没有一致的结论(Meinzer et al., 2017;Henry等 人,2019 年)。研究发现,叶面水分吸收是一种重要的水分获取机制,可以减轻某些树种的水分不足,但叶面水分吸收对气孔调节的作用仍然未知(Eller et al., 2013)。
Structural investment 结构性投资
Stomatal behavior is regulated not only by water potential but also by structural adaptations that impact the demand and supply for water. Structural traits of leaf or stem such as vein density, specific leaf area (SLA), and wood density are essential to the exploring differential stomatal behaviors among species, which show the tradeoff between investment and stress tolerance (Figure 2).
气孔行为不仅受水势的调节,还受影响水供需的结构适应的调节。叶或茎的结构特征,如叶脉密度、比叶面积 (SLA) 和木材密度,对于探索物种间的差异气孔行为至关重要,这表明了投资和抗逆性之间的权衡(图 2)。
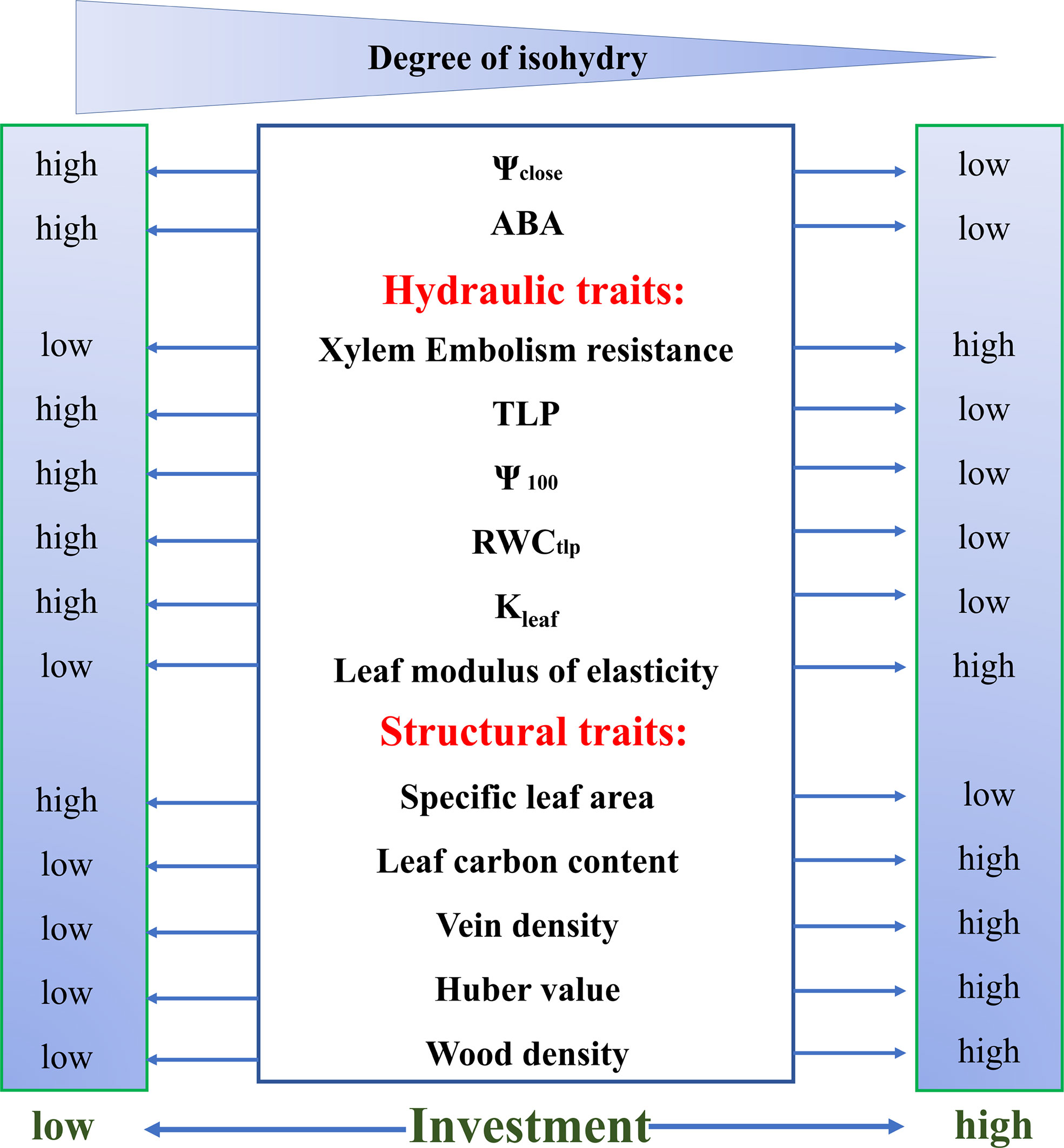
Figure 2 The relationships between the iso/anisohydric behavior and functional traits of leaves and branches, which show how these traits vary with the Ψclose. The anisohydric species show stronger stress resistance, with greater investment in stems and leaves than the isohydric species to maintain stomatal opening under drought conditions. Ψclose (water potential at stomatal closure), TLP (turgor loss point), Ψ100 (osmotic potential at full turgor), RWCtlp (relative water content at TLP), Kleaf (leaf hydraulic conductance). This figure is changed from Chen et al. (2021).
图 2叶和枝的等/非水态行为与功能性状之间的关系,显示了这些性状如何随 Ψ闭合而变化。各水态表现出更强的抗逆性,在干旱条件下维持气孔开放的茎和叶投资比等水态态更大。Ψclose(气孔关闭时的水势)、TLP(膨胀损失点)、Ψ100(完全膨胀时的渗透电位)、RWCtlp(TLP 时的相对含水量)、K叶(叶片水力导率)。这个数字是从 Chen et al. (2021) 改变而来的。
Water flows from the petiole through leaf conduits that form a network of veins (McKown et al., 2010). However, vein ramification is costly in terms of material investment. Vein density was found to be significantly positively correlated with leaf carbon content and the degree of iso/anisohydry, which indicated that anisohydric tree species invested more carbon in leaves than did isohydric species (Chen et al., 2021b).
水从叶柄流过形成叶脉网络的叶导管(McKown et al., 2010)。然而,矿脉分支在材料投资方面成本高昂。发现叶脉密度与叶片碳含量和等/各水态程度呈显著正相关,这表明各水性树种在叶片中投入的碳比等水性树种多(Chen等 ,2021b)。
Specific leaf area (SLA) is the leaf area per unit of dry-mass investment, indicating high SLA requires less investment per unit area. More anisohydric plant species tend to have lower SLA (Pivovaroff et al., 2018; Chen et al., 2021b), and Ψclose was found to be significantly related to leaf carbon content (Chen et al., 2021b). Wood density is an integrative trait and a direct manifestation of investment in wood, which was found to be correlated with the stringency of iso/anisohydry (Klein, 2014; Fu and Meinzer, 2019; Fu et al., 2019). Therefore, it can be concluded that the anisohydric species showed greater investment in foliage and stem than the isohydric species (Chen et al., 2021b; Figure 2).
比叶面积 (SLA) 是每单位干质量投资的叶面积,表明高 SLA 需要较少的单位面积投资。更多的不水植物物种往往具有较低的 SLA(Pivovaroff 等 人,2018 年;Chen et al., 2021b),发现 Ψclose 与叶片碳含量显著相关(Chen et al., 2021b)。木材密度是一种综合特征,是木材投资的直接表现,研究发现它与 iso/anisohydry 的严格性相关(Klein,2014 年;Fu 和 Meinzer,2019 年;Fu et al., 2019)。因此,可以得出结论,非水态物种比等水态物种对叶子和茎的投资更大(Chen等 人,2021b;图 2)。
Variability of stomatal behavior
气孔行为的变异性
Seasonal adjustment in TLP was observed in lianas (Maréchaux et al., 2017), and leaf-level osmotic adjustment was observed in long-term drought experiment (Binks et al., 2016). Adjustments in ϵ have been also documented in the season (Meinzer et al., 2014). A more anisoyhdric species was observed that adjusted TLP during drought, whereas a co-occurring more isohydric species did not (Nolan et al., 2017). Meinzer et al. (2014) observed that the more anisohydric species had more negative TLP and Ψ100 with decreasing water availability, but TLP and Ψ100 in the more isohydric species did not change. In a anisohydric desert shrub, Larrea tridentata, TLP was observed to vary by up to 2 MPa over short periods (Meinzer et al., 1988; Johnson et al., 2018).
在藤本植物中观察到 TLP 的季节性调整 (Maréchaux et al., 2017),在长期干旱实验中观察到叶水平渗透调整 (Binks et al., 2016)。ε的调整也记录在赛季中(Meinzer et al., 2014)。观察到一种更异质的物种在干旱期间调整了 TLP,而同时出现的更等氢的物种则没有(Nolan 等 人,2017 年)。Meinzer 等 人(2014 年)观察到,随着水可用性的降低,非水性更强的物质具有更多的负 TLP 和 Ψ100,但更等水性物质中的 TLP 和 Ψ100 没有变化。在非水性沙漠灌木 Larrea tridentata 中,观察到 TLP 在短时间内变化高达 2 MPa(Meinzer 等 人,1988 年;Johnson et al., 2018)。
Anisohydric species may have to possess the plasticity in leaf P-V traits to compensate that their relatively limited stomatal regulation of maintaining stable leaf water potential (Meinzer et al., 2014; Johnson et al., 2018). In addition to P-V traits, the Ψleaf50 of ansiohydric species also alters during the driest part of the summer (Johnson et al., 2018). Thus, it is reasonable to speculate that anisohydric species may have greater capacities to alter leaf biophysical properties, and individuals of some species may switch between isohydric and anisohydric behavior within species depending on developmental stages and environmental conditions (Meinzer et al., 2014; Meinzer et al., 2016; Johnson et al., 2018).
非水物质可能必须具有叶片 P-V 性状的可塑性,以补偿它们维持稳定叶片水势的相对有限的气孔调节(Meinzer et al., 2014;Johnson et al., 2018)。除了 PV 性状外,水合物物种的 Ψ叶 50 在夏季最干燥的时候也会发生变化(Johnson et al., 2018)。因此,可以合理地推测,各不干物质可能具有更强的改变叶片生物物理特性的能力,并且某些物种的个体可能会根据发育阶段和环境条件在物种内的等水行为和各水行为之间切换(Meinzer et al., 2014;Meinzer et al., 2016;Johnson et al., 2018)。
Xylem embolism resistance
木质部栓塞抵抗
During a paleoclimatic crisis, hydraulic systems evolved under selective pressure to be highly resistant to embolism as increasing drought levels (Larter et al., 2017; Martin-StPaul et al., 2017), and species have evolved broad differences in embolism resistance under this strong selection (Choat et al., 2012).
在古气候危机期间,随着干旱程度的增加,水力系统在选择性压力下进化为对栓塞具有高度抵抗力(Larter 等 人,2017 年;Martin-StPaul等 人,2017 年),并且在这种强选择下物种在栓塞抵抗性方面进化出了广泛的差异(Choat等 人,2012 年)。
Xylem safety threshold is the point in which a tree starts losing control over its hydraulic system. When the water potential is out of the safety threshold, continuing residual water loss can no longer be compensated by water uptake from the soil. Therefore, embolism resistance is one of the key traits responsible for predicting the drought tolerance of tree species (Brodribb and Cochard, 2009; Anderegg et al., 2016; Choat et al., 2018; Brodribb et al., 2020).
赛莱默安全阈值是树木开始失去对其液压系统的控制的点。当水势超出安全阈值时,持续的残余水分损失不能再通过土壤的吸水来补偿。因此,抗栓塞性是负责预测树种耐旱性的关键性状之一(Brodribb 和 Cochard,2009 年;Anderegg et al., 2016;Choat et al ., 2018;Brodribb等 人,2020 年)。
P50 is often used to characterize the xylem safety of trees (Brodribb and Cochard, 2009; Choat et al., 2012). When water potential is below P50, the water transport function will be markedly impaired (Choat et al., 2012; Choat et al., 2018; Arend et al., 2021). The hydraulic system of plants has evolved to the level of embolism resistance reaching P50 down to -18.8 MPa (Larter et al., 2015); and the most vulnerable species is found in tropical forest, whose P50 is as high as -0.18 MPa (Maherali et al., 2004).
P50 通常用于描述树木的木质部安全性(Brodribb 和 Cochard,2009 年;Choat et al., 2012)。当水势低于 P50 时,水的运输功能将明显受损(Choat et et al., 2012;Choat et al ., 2018;Arend et al., 2021)。植物的水力系统已经进化到抗栓塞水平,达到 P50 至 -18.8 MPa(Larter 等人 ,2015 年);最脆弱的物种分布在热带森林中,其 P50 高达 -0.18 MPa(Maherali 等 人,2004 年)。
Angiosperms exhibited higher intraspecific variation than gymnosperms in P50 (Anderegg, 2015). Higher angiosperm variability in P50 ought to result from anatomical differences between gymnosperms and angiosperms (Johnson et al., 2012; Anderegg, 2015). In addition, P50 is not the hydraulic safety threshold of all woody plants. Angiosperm species may be unable to recover if water potential dropped below P88 (the water potential inducing 88% xylem embolism), and P50 is considered more likely to be the hydraulic safety threshold of gymnosperms species (Urli et al., 2013). Environmental conditions and developmental stages may also affect the threshold to some degree. A recent field research of adult Norway spruce showed that processes leading to a rapid deterioration of tree hydraulic status and tree mortality occur in before P50 is reached (Arend et al., 2021; Li et al., 2022), suggesting that the vulnerability of some species to hydraulic failure may differ from previously expectations under different conditions.
在 P50 中,被子植物表现出比裸子植物更高的种内变异(Anderegg,2015)。P50 中较高的被子植物变异性应该是由于裸子植物和被子植物之间的解剖学差异造成的(Johnson et al., 2012;Anderegg,2015 年)。此外,P50 并不是所有木本植物的水力安全阈值。如果水势低于 P88(水势诱导 88% 木质部栓塞),被子植物物种可能无法恢复,并且 P50 被认为更有可能是裸子植物物种的水力安全阈值(Urli 等 人,2013 年)。环境条件和发育阶段也可能在一定程度上影响阈值。最近对挪威云杉成年的实地研究表明,导致树木水力状况迅速恶化和树木死亡率的过程发生在达到 P50 之前(Arend 等 人,2021 年;Li et al., 2022),这表明在不同条件下,某些物种对水力故障的脆弱性可能与以前的预期不同。
The study of xylem embolism has a long history (Dixon and Joly, 1895; Ewart, 1906), and extensive research have been done in this field, including research methods (Cochard et al., 2013; Chen et al., 2021a). Thus, in this review, we do not dwell on the previous common description and only briefly describe the determinants of xylem embolism resistance from the perspective of anatomy.
木质部栓塞的研究由来已久(Dixon 和 Joly,1895 年;Ewart,1906 年),并且在该领域进行了广泛的研究,包括研究方法(Cochard 等 人,2013 年;Chen等 人,2021a)。因此,在这篇综述中,我们不再详述前面的常见描述,只从解剖学的角度简要描述木质部栓塞耐药的决定因素。
The determinants of xylem embolism resistance
木质部栓塞耐药的决定因素
Embolism occurs as ‘air-seeding’ enters the conduits through the pit pores, and pit area hypothesis proposes that the probability of embolism propagation in conduits increases with the total pit area of vessels (Tyree and Zimmermann, 2002; Wheeler et al., 2005). Larger vessels are more likely to have larger total pit area, so larger vessels may be more vulnerable to embolism (Jacobsen et al., 2007). Within species, smaller conduits were found to tend to be less vulnerable to embolism (Cai and Tyree, 2010; Fichot et al., 2015). However, the lack of comprehensive knowledge on the conduit size and pit traits further limits the understanding of the relationship between conduit dimensions and drought-induced embolism resistance (Cardoso et al., 2020). The porosity of pit membranes seems to be more important because pit membrane prevents bubble propagation (Tyree and Zimmermann, 2002). Thus, the importance of conduit size for embolism resistance remains a debate subject.
当“空气种子”通过凹坑孔进入导管时,就会发生栓塞,凹坑面积假说提出,栓塞在导管中传播的可能性随着血管总凹坑面积的增加而增加(Tyree 和 Zimmermann,2002 年;Wheeler et al., 2005)。较大的血管更有可能具有更大的总凹坑面积,因此较大的血管可能更容易发生栓塞(Jacobsen 等 人,2007 年)。在物种内,发现较小的导管往往不易发生栓塞(Cai 和 Tyree,2010 年;Fichot et al., 2015)。然而,缺乏对导管大小和坑特征的全面了解进一步限制了对导管尺寸与干旱诱导的栓塞抵抗力之间关系的理解(Cardoso et al., 2020)。凹坑膜的孔隙率似乎更重要,因为凹坑膜可以防止气泡传播(Tyree 和 Zimmermann,2002 年)。因此,导管大小对栓塞抵抗的重要性仍然是一个争论的话题。
Pit membranes develop from the cell wall made of multiple cellulose layers, pectins and structural proteins, with a thickness between 140 nm and 1180 nm and including between 4 and 30 layers (Fichot et al., 2015; Kaack et al., 2021). The three-dimensional structure, hydration of pit membranes and their chemical composition play a key role in the resistance to embolism spread (Cardoso et al., 2020; Kaack et al., 2021). Perfusion experiments showed that cellulose and pectin are main components of pit membrane determining embolism resistance (Fichot et al., 2015).
凹坑膜由多个纤维素层、果胶和结构蛋白组成的细胞壁发育而来,厚度在 140 nm 和 1180 nm 之间,包括 4 到 30 层(Fichot等 人,2015 年;Kaack et al., 2021)。凹坑膜的三维结构、水合作用及其化学成分在抵抗栓塞扩散中起关键作用(Cardoso等 人,2020 年;Kaack et al., 2021)。灌注实验表明,纤维素和果胶是决定栓塞抵抗性的坑膜的主要成分(Fichot et al., 2015)。
Embolism spreading is related to the porosity of pit membrane, smaller pores tending to increase resistance (Choat et al., 2008; Fichot et al., 2015). Therefore, vulnerability to air-seeding is associated with the pit area, a bigger pit area implying an increased likelihood of having larger pores (Hacke et al., 2006). Thicker pit membrane is related to reduced porosity and therefore increasing resistance to embolism (Fichot et al., 2015; Figure 3). This link between embolism resistance and pit membrane thickness is also valid at the intrageneric (Scholz et al., 2013) and intraspecific level (Schuldt et al., 2016). Variation in pit membrane thickness is largely influenced by the number of microfibril layers, with thin pit membranes consisting of less microfibril layers than thick pit membranes (Kaack et al., 2021).
栓塞扩散与坑膜的孔隙率有关,较小的孔隙往往会增加阻力(Choat et et al., 2008;Fichot et al., 2015)。因此,对空气播种的脆弱性与坑面积有关,更大的坑面积意味着具有更大孔隙的可能性增加(Hacke 等 人,2006 年)。较厚的凹坑膜与孔隙率降低有关,因此增加了对栓塞的抵抗力(Fichot 等 人,2015 年;图 3)。栓塞抵抗与凹坑膜厚度之间的这种联系在属内 (Scholz et al., 2013) 和种内水平 (Schuldt et al., 2016) 也有效。凹坑膜厚度的变化在很大程度上受微纤丝层数量的影响,薄凹坑膜由比厚凹坑膜组成的微纤维层少(Kaack等 人,2021 年)。
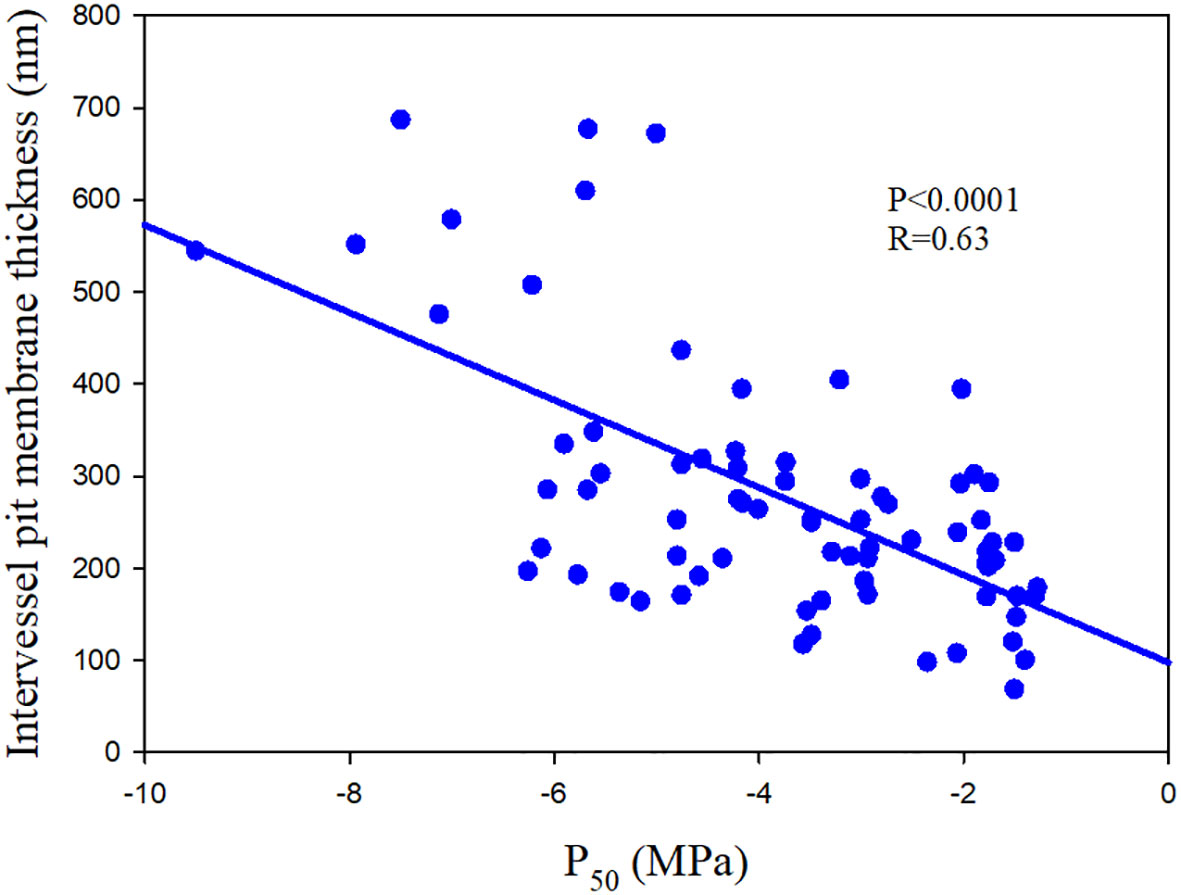
Figure 3 The relationship between water potential at which 50% loss of hydraulic conductance (P50) and intervessel pit membrane thickness of 75 angiosperm speices. The data comes from Scholz et al. (2013); Kaack et al. (2021) and Li et al. (2016).
图 3水势损失 50% 的水力导率 (P50) 与 75 个被子植物的血管间凹坑膜厚度之间的关系。数据来自 Scholz 等 人(2013 年);Kaack 等 人(2021 年)和 Li 等 人(2016 年)。
Species with thicker conduits walls have less porous and thicker pit membranes and consequent stronger embolism resistance (Li et al., 2016). Thick conduit walls also facilitate strong wall reinforcement (conduit thickness/span), which would against deflection and even implosion of conduit walls and possible injury to pit membranes (Hacke and Sperry, 2001; Hacke et al., 2001). An extensive and supportive fiber matrix is also associated with embolism resistance. It was proposed that small diameter fibers can result in dense wood and provide mechanical strength, which facilitate resisting embolism (Jacobsen et al., 2005; Mccullon et al., 2012; Blackman et al., 2019; Chen et al., 2020). Some studies also found that the positive relationship between fiber wall thickness and embolism resistance may be caused by lignification and wood density of xylem (Jacobsen et al., 2005; Awad et al., 2012; Fichot et al., 2015; Torres-Ruiz et al., 2017). Moreover, vasicentric tracheids and fiber–tracheids could act as water reservoirs, thereby increasing tissue capacitance, which are associated with lower cavitation vulnerability (Barotto et al., 2016).
导管壁较厚的物种具有较少的孔隙和较厚的凹坑膜,因此具有更强的栓塞抵抗力(Li et al., 2016)。厚的导管壁还有助于坚固的墙壁加固(导管厚度/跨度),这将防止导管壁的偏转甚至内爆以及可能对坑膜的伤害(Hacke 和 Sperry,2001 年;Hacke et al., 2001)。广泛且支撑性的纤维基质也与栓塞抵抗有关。有人提出,小直径纤维可以产生致密的木材并提供机械强度,从而促进抵抗栓塞(Jacobsen 等 人,2005 年;Mccullon 等 人,2012 年;Blackman et al., 2019;Chen等 人,2020 年)。一些研究还发现,纤维壁厚与栓塞抵抗力之间的正相关可能是由木质部的木质化和木材密度引起的(Jacobsen等 人,2005 年;Awad et al., 2012;Fichot et al., 2015;Torres-Ruiz et al., 2017)。此外,以血管为中心的管胞和纤维管胞可以充当水库,从而增加组织容量,这与较低的空化脆弱性有关(Barotto等 人,2016 年)。
Embolism-resistant traits, such as thick pit membrane, thick conduit wall, high wood density and high fiber-wall area, all are costly in terms of carbon investment, highlighting the relationship between stress resistance and carbon investment (Figure 4).
抗栓塞特性,如厚坑膜、厚导管壁、高木材密度和高纤维壁面积,在碳投资方面都成本高昂,突出了抗应力性和碳投资之间的关系(图 4)。
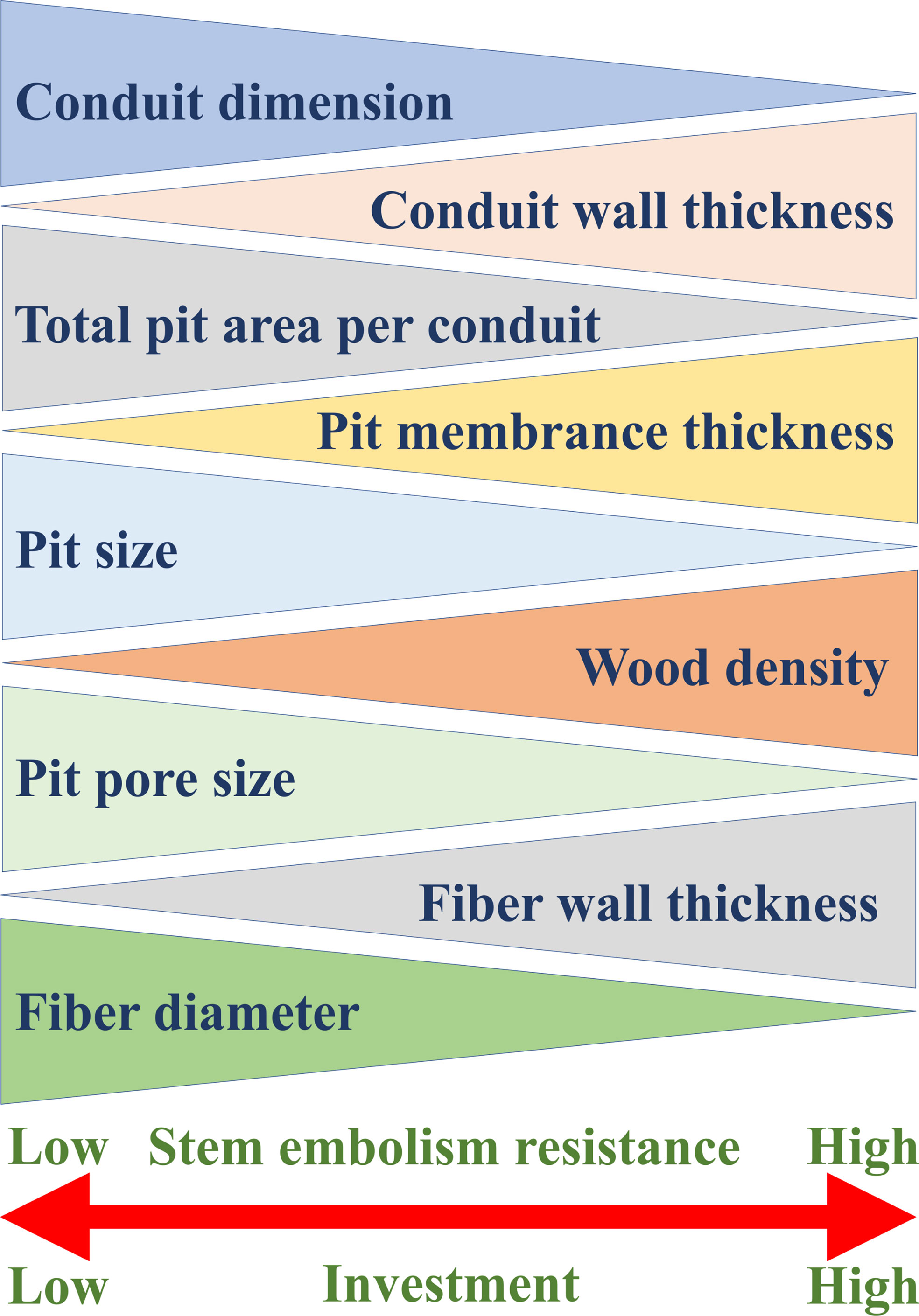
Figure 4 General xylem traits that determine stem embolism resistance and the relationships between these traits and stem embolism resistance. The thicker end of each triangle represents the higher value of the corresponding trait, and the xylem with stronger embolism resistance has greater construction investment.
图 4决定茎栓塞抵抗的一般木质部性状以及这些性状与茎栓塞抵抗性之间的关系。每个三角形较粗的一端代表相应性状的较高值,抗栓塞性较强的木质部具有较大的构建投资。
Variability of embolism resistance
栓塞抵抗的可变性
In general, the intra-specific P50 varied with more negative P50 values found in more xeric environments over space (Anderegg, 2015), and P50 values shifted toward more negative in response to drought stress (Wortemann et al., 2011). However, a few studies revealed that individuals exposed to drought stress were more vulnerable to embolism (Nardini and Luglio, 2014; Savi et al., 2015; Trugman et al., 2021). The explanation of this phenomenon is ‘cavitation fatigue’ for plants experienced chronic or extreme water stress (Hacke et al., 2001; Nardini and Luglio, 2014). Increased vulnerability to cavitation of the species, Quercus ilex L. (holm oak), exposed to long-lasting growing at urban sites under impervious pavements or 6-yr partial rainfall exclusion, might be explained as the hydraulic damage caused by cavitation fatigue (Limousin et al., 2010; Savi et al., 2015). In addition, cavitation fatigue has been also applied to explain the differences of embolism resistance between sapwood and heartwood (Fichot et al., 2015). A previous study demonstrated that conduits of the outer ring were still functional, whereas conduits of the older xylem were almost all embolized (Sperry et al., 1991).
一般来说,种内 P50 变化,在太空上更干燥的环境中发现更多的负 P50 值(Anderegg,2015),而 P50 值在响应干旱胁迫时转向更负值(Wortemann 等 人,2011 年)。然而,一些研究表明,暴露于干旱压力下的个体更容易发生栓塞(Nardini 和 Luglio,2014 年;Savi et al., 2015;Trugman等 人,2021 年)。对这种现象的解释是植物经历过慢性或极端水分胁迫的“空化疲劳”(Hacke et al., 2001;Nardini 和 Luglio,2014 年)。在城市场地,在不透水路面或 6 年部分降雨排除下,栎树 Quercus ilex L.(圣栎)暴露在城市地区长期生长,对空化的影响增加,可以解释为空化疲劳造成的水力损伤(Limousin 等 人,2010 年;Savi et al., 2015)。此外,空化疲劳也被用于解释边材和心材之间抗栓塞性的差异(Fichot et al ., 2015)。先前的研究表明,外环的导管仍然有效,而旧木质部的导管几乎全部栓塞(Sperry et al., 1991)。
The nutritional environment may also affect intra-specific variation of embolism resistance. High N availability tends to increase porosity of the pit membranes of vessel, and therefore, N-fertilized individuals have been shown to be more vulnerable to embolism (Plavcová and Hacke, 2012; Plavcová et al., 2013; Fichot et al., 2015). Additional P supply tends to increase embolism resistance, especially at high N concentrations (Harvey and Van Den Driessche, 1997; Hevia et al., 2019). As the xylem of species/genotypes more resistant to embolism needs more carbon investment and the carbon resources of shaded plants are limited, shaded individuals tend to be more vulnerable to cavitation (Schoonmaker et al., 2010; Plavcová et al., 2011). In addition, phloem conductance and water storage affect the recovery of embolism (Hammond and Adams, 2019). Nevertheless, intra-specific variation of vulnerability to embolism should be small, and a study of one widespread pine species (Pinus pinaster) showed low variation in genes of P50 across different populations (Lamy et al., 2014).
营养环境也可能影响栓塞抵抗性的种内变异。高氮可用性往往会增加血管凹坑膜的孔隙率,因此,氮肥个体已被证明更容易发生栓塞(Plavcová 和 Hacke,2012 年;Plavcová et al., 2013;Fichot et al., 2015)。额外的 P 供应往往会增加栓塞抵抗力,尤其是在高 N 浓度下(Harvey 和 Van Den Driessche,1997 年;Hevia等 人,2019 年)。由于对栓塞更具抵抗力的物种/基因型的木质部需要更多的碳投资,并且遮荫植物的碳资源有限,因此遮荫个体往往更容易受到空化的影响(Schoonmaker等 人,2010 年;Plavcová et al., 2011)。此外,韧皮部电导和水储存会影响栓塞的恢复(Hammond 和 Adams,2019 年)。然而,栓塞易感性的种内变异应该很小,对一种广泛分布的松树种 (Pinus pinaster) 的研究表明,不同种群中 P50 基因的变异较低(Lamy et al., 2014)。
Root properties 根属性
Plant root system plays a significant role in plant growth by exploiting soil resources through the uptake of water and nutrients (Wasaya et al., 2018). In general, resource absorption is undertaken by roots of terminal branch orders (usually the first- and second-order roots), whereas roots of the higher branch orders perform other functions, such as anchorage and storage (McCormack et al., 2015). Trees within the same forest are equipped with different rooting depths, so available water of trees varies with depth. In general, deep roots, i.e. deep-water access, may mitigate drought-induced mortality by limiting exposure to water stress. In a tropical forest, species exposure to drought stress exponentially declined with deeper root depth, which was relevant to drought resistance and resilience (Chitra-Tarak et al., 2021).
植物根系通过吸收水和养分来开发土壤资源,在植物生长中发挥着重要作用(Wasaya et al., 2018)。一般来说,资源吸收是由末端分支目(通常是一级和二级根)的根进行的,而高级分支顺序的根执行其他功能,例如锚定和储存(McCormack等 人,2015)。同一片森林中的树木具有不同的生根深度,因此树木的可用水量随深度而变化。一般来说,深根,即深水通道,可以通过限制暴露于水分胁迫来减轻干旱引起的死亡率。在热带森林中,暴露于干旱胁迫下的物种随着根系深度的加深而呈指数下降,这与抗旱性和恢复力有关(Chitra-Tarak et al., 2021)。
Root traits such as fine root diameter, specific root length, root angle, root length density and root hydraulic traits are considered to be linked with root water acquisition, water use efficiency, drought tolerance and access to nutrient, etc., which are important for understanding plant growth, survival and productivity (Nelson and Oliver, 2017; Wasaya et al., 2018; Tracy et al., 2020). The anatomical traits for the adaptations to soil water content can be determined by indices of the ratio of the root tissue areas (i.e. cortex to stele ratio, xylem to stele ratio) (Yamauchi et al., 2020). Root hairs are subcellular protrusions of the root epidermis, which extend into the soil and can improve the water and nutrient capture (Lynch, 2019). Importantly, root hydraulic traits are directly related to the capabilities and strategies of root organ and even whole plant to adapt to drought. Losing water from the root cells causes dehydrating roots to shrink, physically reduces hydraulic conductance at the soil-root interface, and causes roots to lose turgor which is crucial to root growth (Bartlett et al., 2021b). Roots with a more negative Ψ100 would exhibit a higher cell solute concentration and consequently more negative ΨTLP, and roots with a greater RWCTLP would retain more water at ΨTLP (Bartlett et al., 2021b). A lower root capacitance indicates that the roots retain more volume via less cell shrinkage, and thus maintain greater contact with the soil (Bartlett et al., 2021b). Hydraulic redistribution describes the passive flux of water through roots, for example from moist to dry soil layers (Hafner et al., 2020). Osmotic adjustment would strengthen the water potential gradient driving hydraulic redistribution, and higher hydraulic conductivities and larger conduits are also found to be related to higher hydraulic redistribution quantity (Hafner et al., 2020; Bartlett et al., 2021b). Root cortical lacunae usually precede root xylem embolism under drought stress, and embolism usually appears first in the fine roots and then in older, coarse roots (Cuneo et al., 2016). However, turgor and embolism recovery of roots occurs quickly upon exposure to water (c. 40 min-4 h) by absorbing inorganic ions and water (Shabala and Lew, 2002; Cuneo et al., 2018). Although the research on root hydraulic characteristics has made great progress, the research is still relatively backward compared with the aboveground part. Due to the inaccessibility of roots, new methods for root assaying need to be developed in the future (Nelson and Oliver, 2017).
根性状,如细根直径、比根长、根角、根长密度和根水力性状被认为与根系水分获取、水分利用效率、耐旱性和营养获取等有关,这对于理解植物生长、生存和生产力很重要(Nelson 和 Oliver,2017 年;Wasaya等 人,2018 年;Tracy et al., 2020)。适应土壤含水量的解剖学特征可以通过根组织面积比的指数(即皮层与柱的比率、木质部与柱的比率)来确定(Yamauchi等 人,2020 年)。根毛是根表皮的亚细胞突起,延伸到土壤中,可以改善水分和养分的捕获(Lynch,2019)。重要的是,根系水力特性与根器官甚至整株植物适应干旱的能力和策略直接相关。根细胞中的水分流失会导致脱水的根收缩,物理上降低土壤-根界面的水力传导,并导致根失去对根生长至关重要的膨胀(Bartlett et al., 2021b)。具有较高负 Ψ100 的根将表现出较高的细胞溶质浓度,因此具有更多负 ΨTLP,而具有较高 RWCTLP 的根将在 ΨTLP 处保留更多的水分(Bartlett等 人,2021b)。较低的根电容表明根通过较少的细胞收缩保留更多的体积,从而保持与土壤的更大接触(Bartlett 等 人,2021b)。水力再分配描述了水通过根系的被动通量,例如从潮湿到干燥的土壤层(Hafner 等 人。,2020 年)。渗透调节会加强驱动水力再分配的水势梯度,并且还发现较高的水力传导率和更大的管道与较高的水力再分配量有关(Hafner et al., 2020;Bartlett et al., 2021b)。在干旱胁迫下,根皮质空隙通常先于根木质部栓塞,栓塞通常首先出现在细根中,然后出现在较老的粗根中(Cuneo et al., 2016)。然而,通过吸收无机离子和水,在暴露于水中(约 40 分钟-4 小时)时(约 40 分钟-4 小时),根的膨胀和栓塞恢复迅速发生(Shabala 和 Lew,2002 年;Cuneo et al., 2018)。虽然根系水力特性的研究取得了长足的进步,但与地上部分相比,研究仍然相对落后。由于根的无法接近,未来需要开发新的根测定方法(Nelson 和 Oliver,2017)。
Coordination of stomatal regulation, xylem embolism resistance and root properties
气孔调节、木质部栓塞抵抗力和根特性的协调
The ability of maintaining high surface conductance to CO2 while avoiding desiccation is pivotal for survival in land plants. While reducing leaf Gs for reducing water loss has the cost of constraining photosynthetic carbon sequestration (Cowan and Farquhar, 1977). Therefore, the functional properties of the plant hydraulic system are integral to carbon and water balance (Drake et al., 2015; Figure 5A), and the ability of maximizing carbon gain and minimizing water loss has served as a powerful selective force on the evolution of functional and structural adaptations to drought in plants (Ambrose et al., 2009). Strong correlations between iso-anisohydric behavior and stem traits were observed in recent studies, highlighting the coordination of stomatal regulation and xylem function (Chen et al., 2021b; Figure 6). Many species with similar root syndromes display contrasting aboveground traits, which highlights the importance of including belowground organs to the whole-plant trait integration (Carmona et al., 2021). However, researches on the relationship between stem and leaf traits and root properties are still scarce, thus there is an important research task in the future in order to fully understand the coordination of tree water relations.
在避免干燥的同时保持对 CO2 的高表面电导率的能力对于陆地植物的生存至关重要。虽然减少叶片 Gs 以减少水分流失具有限制光合碳封存的成本(Cowan 和 Farquhar,1977 年)。因此,植物液压系统的功能特性是碳和水平衡不可或缺的一部分(Drake et al., 2015;图 5A),以及最大化碳增益和最小化水分流失的能力已成为植物对干旱的功能和结构适应进化的强大选择力量(Ambrose et al., 2009)。在最近的研究中观察到等水合行为与茎性状之间的强相关性,突出了气孔调节和木质部功能的协调(Chen等人 ,2021b;图 6)。许多具有相似根综合征的物种表现出截然不同的地上性状,这突出了将地下器官纳入全植物性状整合的重要性(Carmona等 人,2021 年)。然而,关于茎叶性状与根性质之间关系的研究仍然很少,因此要充分理解树木水分关系的协调性,未来有一项重要的研究任务。
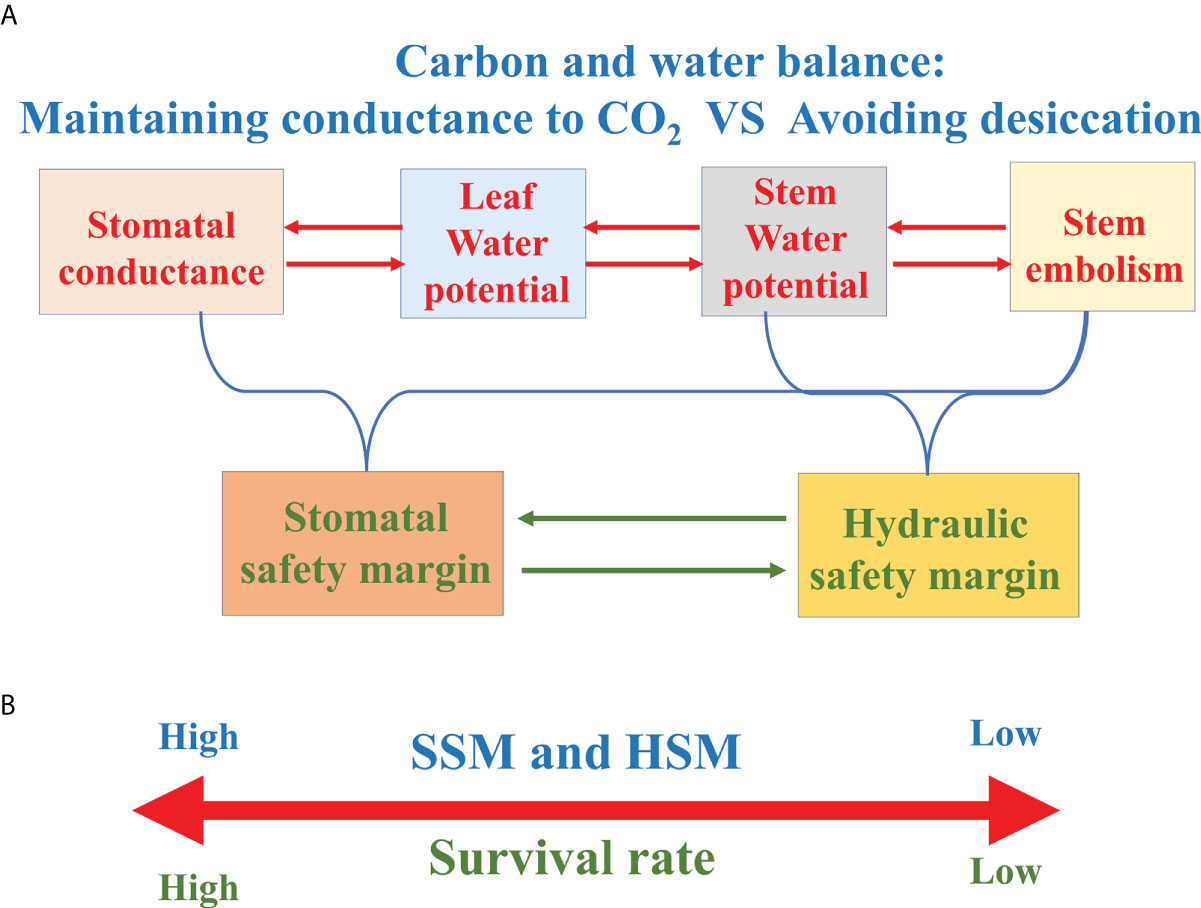
Figure 5 (A) Frame diagram characterizing the relationships between stomatal conductance, leaf/stem water potential and stem xylem embolism. (B) The survival rate of tree species with greater hydraulic safety margin (HSM) or stomatal safety margin (SSM) is higher under drought conditions.
图 5(A) 表征气孔导度、叶/茎水势和茎木质部栓塞之间关系的框架图。(B) 在干旱条件下,具有较大水力安全边际 (HSM) 或气孔安全边际 (SSM) 的树种的成活率更高。
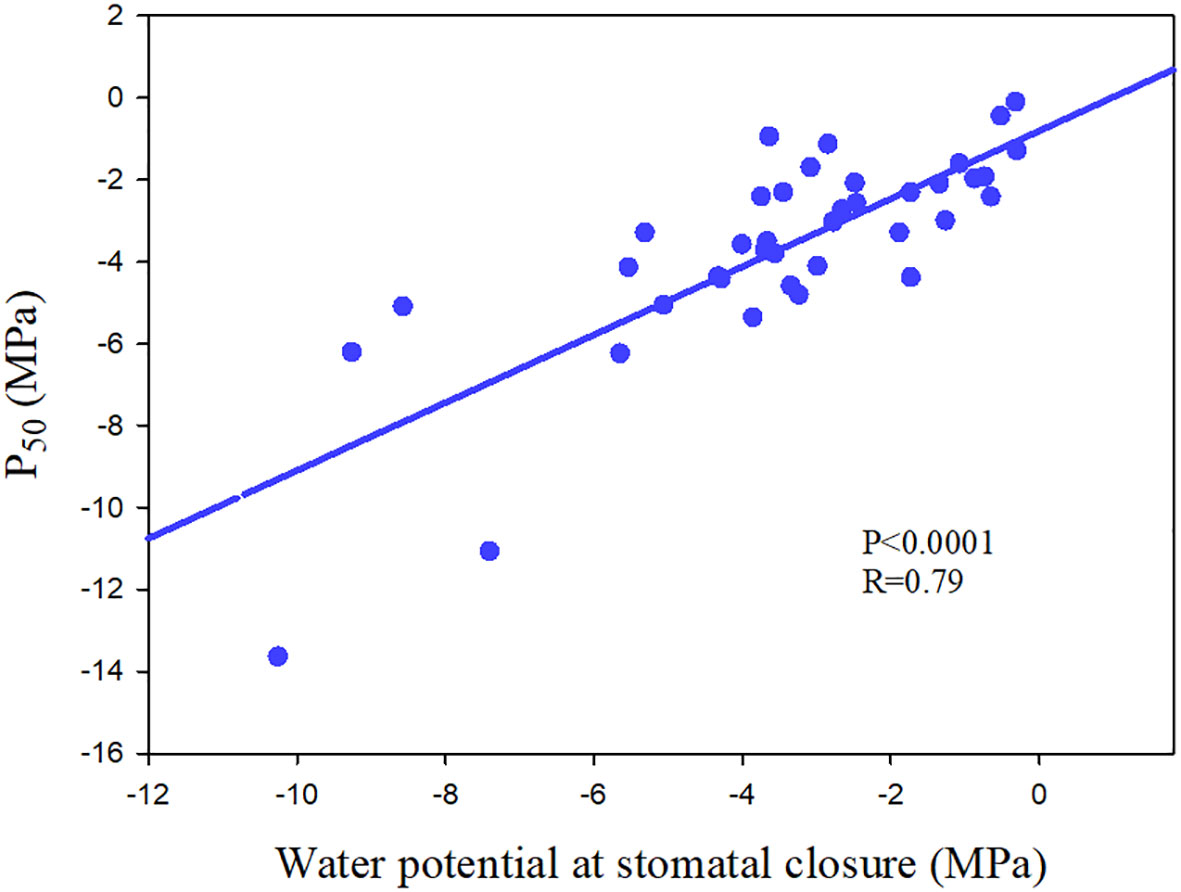
Figure 6 The relationship between water potential at stomatal closure and water potential at which 50% loss of hydraulic conductance (P50) of 39 angiosperm speices. The data comes from Pivovaroff et al. (2018); Chen et al. (2019a) and Li et al. (2015).
图 6气孔闭合时水势与 39 个被子植物特指水力导率损失 50% 的水势 (P50) 之间的关系。数据来自 Pivovaroff 等 人(2018 年);Chen et al. (2019a) 和 Li et al. (2015)。
The correlation of stomatal behavior with xylem properties
气孔行为与木质部特性的相关性
The stomatal closure for preventing Ψleaf from falling below a set minimum value facilitates keeping the stem xylem from experiencing more serious embolism (Meinzer et al., 2014), and xylem embolism may also produce rapid hydraulic signals for initiating the stomatal response (Salleo et al., 2000). The hydraulic message triggered by embolism including ABA and pH changes in the xylem sap that travels to the shoots and contributes to the regulation of Gs, together with the direct effect of xylem water potential (Salleo et al., 2000; Tardieu, 2016). In other words, stomatal regulation and stem embolism are mutually linked by a negative feedback mechanism (Salleo et al., 2000).
用于防止 Ψ叶低于设定最小值的气孔闭合有助于防止茎木质部发生更严重的栓塞(Meinzer et al., 2014),木质部栓塞也可能产生快速的液压信号来启动气孔反应(Salleo et al., 2000)。由栓塞触发的水力信息包括木质部汁液中的 ABA 和 pH 值变化,该汁液传播到嫩芽并有助于调节 Gs,以及木质部水势的直接影响(Salleo 等 人,2000 年;Tardieu,2016 年)。换句话说,气孔调节和茎栓塞通过负反馈机制相互联系(Salleo et al., 2000)。
The iso/anisohydric strategies combined with the corresponding xylem traits reflect the drought avoidance strategy and drought tolerance strategy. Drought avoidance and drought tolerance are two divergent strategies promoting plants adaptation to drought (Oliveira et al., 2021). The continuum of stomatal strategy from isohydry to anisohydric basically coincides with that from drought avoidance to drought tolerance. Drought-avoiding species close stomata rapidly responding to increased water deficit, and even directly shed leaves, which constrains carbon assimilation during dry periods; such drought avoiders with low wood density and high hydraulic capacitance appear to avoid xylem embolism via releasing of stored water to decrease Ψleaf fluctuations (Brodribb et al., 2014; Oliveira et al., 2021). On the contrary, species of drought tolerance with denser wood and low capacitance appear to rely on xylem structure to resist embolism and could maintain foliage over dry periods (Fu et al., 2019). Therefore, drought tolerators can sustain relatively photosynthesis, albeit at the cost of enhanced investment in structural reinforcement of embolism-resistant xylem and in osmoregulation to sustain leaf turgor (Meinzer et al., 2016). In general, anisohydric species appear to occupy more drought-prone habitats compared with isohydric species (McDowell et al., 2008).
iso/isosohydric 策略与相应的木质部性状相结合,反映了避旱策略和耐旱策略。避免干旱和耐旱是促进植物适应干旱的两种不同的策略(Oliveira et al., 2021)。气孔策略从等水态到非水态的连续体与从避旱到耐旱的连续体基本一致。避免干旱的物种靠近气孔,对增加的水分短缺做出快速反应,甚至直接脱落叶子,这限制了干旱时期的碳同化;这种具有低木材密度和高水力电容的干旱避免剂似乎通过释放储存的水来减少 Ψ叶的波动来避免木质部栓塞(Brodribb等 人,2014 年;Oliveira等 人,2021 年)。相反,具有更致密木材和低容量的耐旱物种似乎依靠木质部结构来抵抗栓塞,并且可以在干燥期间保持叶子(Fu et al., 2019)。因此,耐旱剂可以维持相对的光合作用,尽管代价是加强了对抗栓塞木质部结构加固和渗透调节以维持叶片膨胀的投资(Meinzer et al., 2016)。一般来说,与等水物质相比,各水物质似乎占据了更易发生干旱的栖息地(McDowell 等 ,2008)。
Hydraulic safety margin 液压安全裕度
The concept of HSM is generally described as the difference between a critical point (usually P50 or P88) on the vulnerability curve to embolism and the minimum seasonal xylem water potential (Ψmin). In view of the significant correlation between the Ψmin and Ψclose, Ψmin is often assumed to provide a measure of stomatal regulation of water potential (Pivovaroff et al., 2018; Chen et al., 2019a). As woody plants tend to maximize carbon sequestration relative to the investments of constructing or maintaining hydraulic support for foliage, woody species routinely operate at the catastrophic brink of xylem failure, highlighting the importance of the margin between xylem water potential and P50 or P88 (Tyree and Sperry, 1988; Bond and Kavanagh, 1999). Therefore, large positive HSM implies a relatively conservative response and small HSM (or even negative HSM) suggests a hydraulically risky response in the control of plant water relations (Skelton et al., 2015; Trugman et al., 2021). Although drought-induced tree death is complex and involves many physiological traits and processes, numerous previous studies showed that HSM was good (even only) predictor of mortality or branch dieback for tree species or shrub species (Adams et al., 2017; Chen et al., 2019a; Figure 5B).
HSM 的概念通常被描述为栓塞脆弱性曲线上的临界点(通常为 P50 或 P88)与最小季节性木质部水势 (Ψmin) 之间的差值。鉴于 Ψmin 和 Ψclose 之间的显著相关性,通常假设 Ψmin 提供了水势气孔调节的量度(Pivovaroff等 人,2018 年;Chen等 人,2019a)。由于木本植物相对于建造或维持叶子水力支撑的投资而言,往往能最大限度地实现碳封存,因此木本植物经常在木质部失效的灾难性边缘运行,突出了木质部水势与 P50 或 P88 之间边缘的重要性(Tyree 和 Sperry,1988 年;Bond 和 Kavanagh,1999 年)。因此,大的正 HSM 意味着相对保守的反应,而小的 HSM(甚至负 HSM)表明在控制植物水关系方面存在水力风险反应(Skelton 等 人,2015 年;Trugman等 人,2021 年)。尽管干旱引起的树木死亡很复杂,涉及许多生理特征和过程,但之前的大量研究表明,HSM 是树种或灌木物种死亡率或树枝枯死的良好(甚至唯一)预测指标(Adams 等 人,2017 年;Chen等 人,2019a;图 5B)。
According to the definition of HSM, HSM is determined by both Ψmin and embolism resistance. Although embolism resistance is strongly associated with the Ψmin across species, more embolism-resistant species have larger hydraulic safety margin than sensitive ones (Markesteijn and Poorter, 2011; Fu and Meinzer, 2019). HSM also affects species-specific distribution, and species that inhabit more arid environments usually possess larger HSM than those occupying wetter ones (Garzón et al., 2018). In addition, there appears to be a tendency for coniferous species to have larger HSMs compared with most angiosperm species (Choat et al., 2012), with Mediterranean climate angiosperms being a potential exception (Meinzer and McCulloh, 2013). Pioneer species usually operate lower HSM than shade-tolerant species which usually have more conservative strategies, and strong differences in HSM were also found between co-occurring deciduous and evergreen species (Markesteijn and Poorter, 2011). To offset a shorter growing season, deciduous species generally maximize photosynthetic carbon intake in the growth season (Wright et al., 2004; Ishida et al., 2006), and would thus operate a greater danger in hydraulic failure to maximize productivity in short growth season (Markesteijn and Poorter, 2011).
根据 HSM 的定义,HSM 由 Ψmin 和栓塞抵抗共同决定。尽管栓塞抵抗与物种之间的 Ψmin 密切相关,但更多的栓塞抵抗物种比敏感物种具有更大的液压安全边际(Markesteijn 和 Poorter,2011 年;Fu 和 Meinzer,2019 年)。HSM 还影响物种特异性分布,栖息在更干旱环境中的物种通常比占据较潮湿环境的物种具有更大的 HSM(Garzón 等 人,2018 年)。此外,与大多数被子植物物种相比,针叶树物种似乎倾向于具有更大的 HSM(Choat 等 人,2012 年),地中海气候被子植物是一个潜在的例外(Meinzer 和 McCulloh,2013 年)。先锋物种通常比耐阴物种的 HSM 更低,后者通常具有更保守的策略,并且在同时出现的落叶树种和常绿树种之间也发现了 HSM 的强烈差异(Markesteijn 和 Poorter,2011)。为了抵消较短的生长季节,落叶树种通常会在生长季节最大限度地吸收光合碳(Wright et al., 2004;Ishida et al., 2006),因此在水力故障中会带来更大的危险,无法在短时间内最大限度地提高生产力(Markesteijn 和 Poorter,2011)。
Unfortunately, for acquiring HSM, datasets of Ψmin in field conditions are typically patchy because of the laborious onsite measurement techniques (Breshears et al., 2009; Choat et al., 2018), and long-term measurement of Ψmin across seasonal scales is very time consuming. In addition, the hydraulic resistance in leaves is dynamic over a day timescale, which would lead to pronounced transpiration-induced disequilibrium between leaf and stem water potential in transpiring shoots (Meinzer et al., 2009). Therefore, stem xylem water potential of field grown plants must be measured on non-transpiring (shading treatment) leaves or shoot tips attached to transpiring shoots (Meinzer et al., 2009), which also increases the difficulty of measuring the Ψmin especially in tall trees.
不幸的是,对于获取 HSM,由于费力的现场测量技术,现场条件下 Ψmin 的数据集通常是不完整的(Breshears等 人,2009 年;Choat等 人,2018 年),并且跨季节尺度长期测量 Ψmin 非常耗时。此外,叶子中的水力阻力在一天的时间尺度上是动态的,这将导致蒸腾作用诱导的蒸腾作用在蒸腾芽中的叶子和茎之间的水势明显不平衡(Meinzer et al., 2009)。因此,必须在非蒸腾(遮荫处理)叶子或附着在蒸腾芽上的芽尖上测量田间植物的茎木质部水势(Meinzer et al., 2009),这也增加了测量 Ψmin 的难度,尤其是在高大的树木中。
Stomatal safety margin 气孔安全边际
Stomatal safety margin (SSM) refers to the difference between Ψclose and the water potential causing xylem dysfunction (usually P50). Compared with HSM, SSM uses Ψclose instead of Ψmin, which offsets the difficulty of obtaining the Ψmin data mentioned above. In addition, SSM more directly integrates the “safety” of stomatal response to water potential and xylem to drought-induced embolism (Skelton et al., 2015; Figure 5A).
气孔安全边际 (SSM) 是指 Ψclose 与导致木质部功能障碍的水势 (通常为 P50) 之间的差异。与 HSM 相比,SSM 使用 Ψclose 而不是 Ψmin,这抵消了获取上述 Ψmin 数据的难度。此外,SSM 更直接地整合了气孔对水势的反应和木质部对干旱诱导的栓塞的“安全性”(Skelton 等 人,2015 年;图 5A)。
Stomatal regulation preventing leaf water potential from falling below a set minimum value facilitates constraining excessive loss of stem hydraulic conductivity. This is in agreement with the hydraulic segmentation hypothesis, i.e., more distal components such as foliage protect stems by earlier embolism under drought (Tyree and Ewers, 1991; Nolf et al., 2015). Large positive SSM represent that leaves stomatal closure occurs before stem severe embolism, whereas negative SSM indicate stomatal closure subsequent to P50 (Skelton et al., 2015).
气孔调节防止叶片水分潜力低于设定的最小值,有助于限制茎水力传导率的过度损失。这与水力分割假说一致,即在干旱下,更远端的成分(如叶子)通过早期栓塞保护茎(Tyree 和 Ewers,1991 年;Nolf et al., 2015)。大的阳性 SSM 表示叶气孔关闭发生在茎严重栓塞之前,而负 SSM 表示气孔在 P50 之后关闭(Skelton et al., 2015)。
Ψclose varied from -0.655 MPa to -5.54 MPa (Martin-StPaul et al., 2017; Chen et al., 2019a), with the outlier about -10 MPa in a chaparral species in Mediterranean-type climate (Pivovaroff et al., 2018), spanning a range of variation about one-third that for embolism resistance (P50, -0.18 MPa to -18.8 MPa) (Maherali et al., 2004; Larter et al., 2015). Meta-analysis showed that most species have Ψclose values that are higher than their P50 (negative value) (Martin-StPaul et al., 2017; Figure 6). However, in the published data, it is found that the stomatal closure of a little tree species occurs after the occurrence of large xylem embolism, and such outliers are easy to die during drought (Skelton et al., 2015; Pivovaroff et al., 2018; Chen et al., 2019a; Chen et al., 2019b). In addition, there are species with strong embolism-resistant stems ultimately experience high mortality, which may result from the terrible coordination between stomatal regulation and xylem embolism resistance (Anderegg et al., 2019). There are clear evidences of partial and complete mortality correlated with hydraulic failure in both isohydric and anisohydric plants (McDowell, 2011), which further highlight the importance of coordination between stomatal regulation and xylem embolism resistance.
Ψclose 从 -0.655 MPa 到 -5.54 MPa 不等(Martin-StPaul et al., 2017;Chen et al., 2019a),在地中海型气候的 chaparral 物种中,异常值约为 -10 MPa(Pivovaroff et al., 2018),跨越的变化范围约为栓塞抵抗性的三分之一(P50,-0.18 MPa 至 -18.8 MPa)(Maherali et al., 2004;Larter et al., 2015)。荟萃分析表明,大多数物种的 Ψ接近值高于其 P50(负值)(Martin-StPaul et al., 2017;图 6)。然而,在已发表的数据中,发现一个小树种的气孔关闭发生在大木质部栓塞发生之后,并且这样的异常值在干旱期间很容易死亡(Skelton et al., 2015;Pivovaroff等 人,2018 年;Chen等 人,2019a;Chen等 人,2019b)。此外,有些具有强栓塞抗性茎的物种最终会经历高死亡率,这可能是由于气孔调节和木质部栓塞耐药性之间的可怕协调造成的(Anderegg等 人,2019 年)。有明确的证据表明,等水态和各水态工厂的部分和完全死亡与水力故障相关(McDowell,2011 年),这进一步强调了气孔调节和木质部栓塞抵抗性之间协调的重要性。
After stomatal closure, it still shows a decline of water potential, which is likely driven by cuticular and residual stomatal conductance. Nonetheless, the time of desiccation of species with wider SSM was longer (Blackman et al., 2019; Cardoso et al., 2020), and species that closing stomata relatively late also assume risky safety margin (Skelton et al., 2015). In general, SSM increased continuously with increasing embolism resistance (Martin-StPaul et al., 2017), and SSM is correlated with the HSM (Chen et al., 2019a). Most importantly, merging stomatal regulation strategies with xylem embolism resistance strategies contributes to a more comprehensive framework to manifest plant adaptation to drought (Skelton et al., 2015).
气孔关闭后,它仍然显示出水势下降,这可能是由表皮和残余气孔导度驱动的。尽管如此,具有更宽 SSM 的物种的干燥时间更长(Blackman等 人,2019 年;Cardoso等人 ,2020 年),并且相对较晚关闭气孔的物种也承担了风险安全边际(Skelton等 人,2015 年)。一般来说,SSM 随着栓塞抵抗力的增加而持续增加(Martin-StPaul等 人,2017 年),并且 SSM 与 HSM 相关(Chen等 人,2019a)。最重要的是,将气孔调节策略与木质部栓塞抵抗策略相结合,有助于形成一个更全面的框架来表现植物对干旱的适应(Skelton等 人,2015 年)。
Coordination between leaf and stem xylem traits and root properties
叶和茎木质部性状与根特性之间的协调
The dialogue between leaves, stems and roots is important in water uptake, transport and utilization. Water uptake occurs when the water potential of root system is higher than leaf water potential, thus establishing a gradient for water flux. In general, there is a linkage between the circadian oscillations in root hydraulic conductance and the daily variations in the transpiration, with significant consequences on water uptake (Tardieu et al., 2017; Trugman et al., 2021).
叶、茎和根之间的对话在水分的吸收、运输和利用中很重要。当根系的水势高于叶片的水势时,就会发生水分吸收,从而建立水通量梯度。一般来说,根系水力传导的昼夜节律振荡与蒸腾作用的每日变化之间存在联系,对水分吸收有重大影响(Tardieu et al., 2017;Trugman等 人,2021 年)。
In terms of coordination between roots and leaf stomata, varieties in soil-plant hydraulic conductance would drive stomatal closure (Cai et al., 2022). Root-produced signaling may also affect stomatal conductance, and split root experiments have the potential to enable the investigation of the water uptake pattern of root systems (Koebernick et al., 2015). Previous studies found that when the dry part of the root system is at an intermediate soil water status, signal output from the entire root system is maximized, such that further soil drying actually decreases root-to-shoot signaling (Dodd et al., 2010). Alternatively, root capacitance might also impact gas exchange. Roots with a lower capacitance, which indicates that roots retain greater volume as water potentials decline, tend to maintain greater gas exchange through smaller declines in root volume and less ABA production (Bartlett et al., 2021b).
在根和叶气孔之间的协调方面,土壤-植物水力导度的多样性将驱动气孔关闭(Cai et al., 2022)。根产生的信号传导也可能影响气孔导度,分根实验有可能研究根系的水分吸收模式(Koebernick et al., 2015)。以前的研究发现,当根系的干燥部分处于中等土壤水分状态时,整个根系的信号输出最大化,因此进一步的土壤干燥实际上降低了根-芽信号传导(Dodd et al., 2010)。或者,根电容也可能影响气体交换。电容较低的根,这表明随着水势的下降,根保持更大的体积,往往通过根体积的较小下降和较少的 ABA 产生来保持更大的气体交换(Bartlett 等 人,2021b)。
In terms of coordination between roots and stem hydraulic traits, there may be a trade-off between root depth and stem xylem embolism resistance. A study found that species exposure to drought stress declined with deeper root depth indicating that trees compensate for drought stress-related mortality risk through deep-water access, whereas species with deeper root had lower stem xylem embolism resistance and narrower stem hydraulic safety margins (Chitra-Tarak et al., 2021; Trugman et al., 2021). Species with investment in deep roots can access reliable deep-water resources, which ensures that hydraulic risk may not realize for them. Nevertheless, shallow-rooted species may tend to pay the cost of stem hydraulic safety, adapting for an environment in which hydraulic risk may be prone to occur, as extreme droughts usually occur in shallow soil layers (Chitra-Tarak et al., 2021). In addition, root systems have metabolic costs of respiration, another trade-off therefore may exist between the carbon cost of the root system and the growth of other organs (for example, stem) (Tardieu et al., 2017).
在根和茎水力性状之间的协调方面,根深和茎木质部栓塞阻力之间可能存在权衡。一项研究发现,物种暴露于干旱胁迫下的程度随着根系深度的降低而下降,这表明树木通过深水通道补偿了与干旱胁迫相关的死亡风险,而根系较深的物种具有较低的茎木质部栓塞抵抗力和较窄的茎液压安全边际(Chitra-Tarak et al., 2021;Trugman等 人,2021 年)。投资于深根的物种可以获得可靠的深水资源,这确保了它们可能无法意识到水力风险。然而,浅根物种可能倾向于支付茎液压安全的成本,以适应可能容易发生液压风险的环境,因为极端干旱通常发生在浅层土壤(Chitra-Tarak 等 人,2021 年)。此外,根系具有呼吸的代谢成本,因此根系的碳成本与其他器官(例如茎)的生长之间可能存在另一种权衡(Tardieu et al., 2017)。
Perspectives 观点
As mentioned above, sound knowledge of tree hydraulic function is crucial to understand drought-induced tree mortality, and it is necessary to extend and integrate other functional traits and processes to establish a more comprehensive framework.
如上所述,对树木水力功能的充分了解对于理解干旱引起的树木死亡至关重要,并且有必要扩展和整合其他功能性状和过程以建立一个更全面的框架。
Stomatal regulation and xylem embolism resistance are likely to interact with other important traits and processes, e.g., leaf area, plant water storage, root properties, hydraulic segmentation, minimum bark conductance and metabolic status, in the plant responses to drought (Hammond and Adams, 2019; Trugman et al., 2021). We can take stomatal regulation and xylem embolism resistance as the core, expand other important traits, including the relationship between investment of matter allocation and stress resistance, and incorporate the thoughts of the ‘fast-slow’ plant economics spectrum to establish a more comprehensive combination of traits that reflects tree species’ response and adaptation to drought. In functionally diverse communities, an integrated traits combination may produce more diverse drought adaptation strategies (Martínez-Vilalta and Garcia-Forner, 2017) and will accurately improve our prediction of how plants to adapt to spatio-temporal changes in environment conditions, which facilitates to model drought-related tree mortality and changes in plant demography in the future (Rowland et al., 2021).
气孔调节和木质部栓塞抵抗性可能与其他重要性状和过程相互作用,例如,在植物对干旱的反应中,叶面积、植物储水、根系特性、水力分割、最小树皮导率和代谢状态(Hammond 和 Adams,2019 年;Trugman等 人,2021 年)。我们可以以气孔调控和木质部栓塞抗性为核心,扩展其他重要性状,包括物质分配投入与抗逆性之间的关系,并结合“快-慢”植物经济学谱系的思想,建立更全面的反映树种对干旱的反应和适应的性状组合。在功能多样化的群落中,综合性状组合可能会产生更多样化的干旱适应策略(Martínez-Vilalta 和 Garcia-Forner,2017),并将准确改进我们对植物如何适应环境条件时空变化的预测,这有助于模拟与干旱相关的树木死亡率和未来植物种群变化(Rowland et al., 2021)。
In addition, we propose four specific urgent issues as future research priorities:
此外,我们提出了四个具体的紧迫问题作为未来的研究重点:
(1) We should expand future works to not only quantify empirical thresholds for hydraulic failure and associated HSM or SSM but also understand species specific variations over time to reach the thresholds (P50, Ψclose, and so on), which requires further knowledge of a range of other traits such as rooting depth, phloem conductance and plant water storage, and so on (Hammond and Adams, 2019).
(1) 我们应该扩大未来的工作,不仅要量化水力故障和相关的 HSM 或 SSM 的经验阈值,还要了解物种随时间的特定变化以达到阈值(P50、Ψclose 等),这需要进一步了解一系列其他特征,例如生根深度、韧皮部电导率和植物储水。 等等(Hammond 和 Adams,2019 年)。
(2) There is uncertainty in the mechanisms about the refilling of embolized xylem and the point of no return along P50, Ψclose, HSM or SSM, which limits our ability to understand and predict the legacies of successive drought events. This uncertainty is also related to the lack of knowledge about the interdependence of hydraulic failure and carbon starvation. Increased embolism and associated xylem tensions would cause declining carbon gain, transport, utilization, and subsequent feedbacks by which declining NSC availability impacts embolism through refilling or water homeostasis, but studies in this area is still lacking (McDowell, 2011; Hammond and Adams, 2019).
(2) 栓塞木质部再填充机制和沿 P50、Ψclose、HSM 或 SSM 的不归点存在不确定性,这限制了我们理解和预测连续干旱事件遗留问题的能力。这种不确定性还与缺乏关于水力故障和碳饥饿相互依存关系的知识有关。栓塞和相关的木质部张力增加会导致碳增益、运输、利用和随后的反馈下降,其中 NSC 可用性的下降通过再填充或水稳态影响栓塞,但该领域的研究仍然缺乏(McDowell,2011 年;Hammond 和 Adams,2019 年)。
(3) It is not fully understood how iso/anisohydric behavior, xylem embolism resistance and HSM or SSM are affected by the root elements, especially root depth and root capacitance. Tree species with different iso/anisohydric behaviors and HSM or SSM may have different investments in above-ground woody stem and below-ground root growth (Ambrose et al., 2015). In addition, it is not clear that how chemical messages originating from roots affect canopy stomatal regulation under field conditions, especially for tall trees (Tardieu, 2016), and how roots as storage organs compensate for aboveground death by resprouting.
(3) 目前尚不完全了解等/各水合行为、木质部栓塞阻力和 HSM 或 SSM 如何受到根元件的影响,尤其是根深和根电容。具有不同等/各水力学行为和 HSM 或 SSM 的树种可能对地上木质茎和地下根系生长有不同的投资(Ambrose et al., 2015)。此外,尚不清楚源自根的化学信息在田间条件下如何影响树冠气孔调节,尤其是对于高大的树木(Tardieu,2016),以及根作为储存器官如何通过发芽来补偿地上死亡。
(4) The increase of the air temperature is the main hallmark of global climate change and is coupled with droughts, hence directly or indirectly affecting drought adaptation/resistance/resilience strategies of tree species. Some species exhibited higher stomatal conductance and stomatal density in the wet season in warming experiment (Wu et al., 2020). However, there is a lack of research on the effect of long-term warming on iso/anisohydric regulation, stem xylem embolism resistance and HSM or SSM, and thus future research in this area should be carried out.
(4) 气温升高是全球气候变化的主要标志,与干旱相辅相成,从而直接或间接影响树种的干旱适应/抵抗/恢复策略。在变暖实验中,一些物种在雨季表现出较高的气孔导度和气孔密度 (Wu et al., 2020)。然而,目前缺乏关于长期增温对 iso/anisohydration 调节、茎木质部栓塞抵抗和 HSM 或 SSM 影响的研究,因此应进一步开展该领域的研究。
Author contributions 作者贡献
ZC, XW and SRL conceived the ideas. ZC, SL, and SRL led the writing of the manuscript. ZC and XW revised the manuscript. All authors contributed to the article and gave final approval for publication.
ZC、XW 和 SRL 构思了这些想法。ZC、SL 和 SRL 领导了手稿的撰写。ZC 和 XW 修改了手稿。所有作者都为本文做出了贡献,并最终批准了出版。
Funding 资金
The research was supported by the Fundamental Research Funds of Chinese Academy of Forestry (CAFYBB2020QB009), the Ministry of Science and Technology of China for Key R&D Program (2021YFD2200405) and the National Natural Science Foundation of China (31800513).
该研究得到了中国林业科学研究院基本科研业务费资金(CAFYBB2020QB009)、中国科学技术部重点研发计划(2021YFD2200405)和国家自然科学基金(31800513)的支持。
Acknowledgments 确认
We would like to thank Baotianman Forest Ecosystem Research Station for the assistance in the previous research related to the ideas of this paper.
感谢宝天满森林生态系统研究站对本文思想相关前期研究的帮助。
Conflict of interest 利益冲突
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
作者声明,该研究是在没有任何可能被解释为潜在利益冲突的商业或财务关系的情况下进行的。
Publisher’s note 出版商注
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
本文中表达的所有声明仅代表作者的观点,并不一定代表其附属组织或出版商、编辑和审稿人的观点。本文中可能评估的任何商品或制造商可能提出的声明均不受出版商的保证或认可。
Supplementary material 补充材料
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.926535/full#supplementary-material
本文的补充材料可在线找到:https://www.frontiersin.org/articles/10.3389/fpls.2022.926535/full#supplementary-material
References 引用
Adams, H. D., Zeppel, M. J.B., Anderegg, W. R.L., Landhäusser, S. M., Tissue, D. T. (2017). A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat Ecol Evol. 1, 1285–1291.
亚当斯,HD,齐佩尔,MJAB,安德雷格,WRL,兰德豪瑟,SM,组织,DT(2017 年)。干旱诱导树木死亡的生理机制的多物种综合。Nat Ecol Evol.1, 1285–1291.
Ambrose, A. R., Baxter, W. L., Wong, C. S., Næsborg, R. R., Williams, C. B., Dawson, T. E. (2015). Contrasting drought-response strategies in California redwoods. Tree Physiol. 35, 453–469. doi: 10.1093/treephys/tpv016
安布罗斯,AR,巴克斯特,WL,黄,CS,奈斯堡,RR,威廉姆斯,CB,道森,TE(2015 年)。加州红杉的干旱应对策略对比。树木生理学。35, 453–469.doi: 10.1093/treephys/TPV016
PubMed Abstract | CrossRef Full Text | Google Scholar
PubMed 摘要 |CrossRef 全文 |谷歌学术
Ambrose, A. R., Sillett, S. C., Dawson, T. E. (2009). Effects of tree height on branch hydraulic, leaf structure and gas exchange in California redwood. Plant Cell Environ. 32, 743–757. doi: 10.1111/j.1365-3040.2009.01950.x
安布罗斯,AR,西莱特,SC,道森,TE(2009 年)。树高对加州红木树枝水力、叶片结构和气体交换的影响。植物细胞环境。32, 743–757.doi: 10.1111/j.1365-3040.2009.01950.x
PubMed Abstract | CrossRef Full Text | Google Scholar
PubMed 摘要 |CrossRef 全文 |谷歌学术
Anderegg, W. R. L. (2015). Spatial and temporal variation in plant hydraulic traits and their relevance for climate change impacts on vegetation. New Phytol. 205, 1008–1014. doi: 10.1111/nph.12907
安德雷格,WRL(2015 年)。植物水力性状的时空变化及其与气候变化对植被影响的相关性。新植物醇。205, 1008–1014.doi: 10.1111/nph.12907
Anderegg, W. R. L., Klein, T., Bartlett, M., Sack, L., Pellegrini, A. F. A., Choat, B., et al. (2016). Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced treemortality across the globe. Proc. Natl. Acad. Sci. U.S.A. 113), 5024–5029. doi: 10.1073/pnas.1525678113
Anderegg, W. R. L., Trugman, A. T., Bowling, D. R., Salvucci., G., Tuttle, S. E. (2019). Plant functional traits and climate influence drought intensification and land–atmosphere feedbacks. Proc. Natl. Acad. Sci. U.S.A. 116, 14071–14076. doi: 10.1073/pnas.1904747116
Arend, M., Link, R. M., Patthey, R., Hoch, G., Schuldt, B., Kahmen, A. (2021). Rapid hydraulic collapse as cause of drought-induced mortality in conifers. Proc. Natl. Acad. Sci. U.S.A. 118, e2025251118. doi: 10.1073/pnas.2025251118
Awad, H., Herbette, S., Brunel, N., Tixier, A., Pilate, G., Cochard, H., et al. (2012). No trade-off between hydraulic and mechanical properties in several transgenic poplars modified for lignins metabolism. Environ. Exp. Bot. 77, 185–195. doi: 10.1016/j.envexpbot.2011.11.023
Barnes, C. R. (1904). The ascent of water in trees. Science. 20, 179–179. doi: 10.1126/science.20.501.179.b
Barotto, A. J., Fernandez, M. E., Gyenge, J., Meyra, A., Martinez-Meier, A., Monteoliva, S. (2016). First insights into the functional role of vasicentric tracheids and parenchyma in eucalyptus species with solitary vessels: Do they contribute to xylem efficiency or safety? Tree Physiol. 36, 1485–1497. doi: 10.1093/treephys/tpw072
Bartlett, M. K., Klein, T., Jansen, S., Choat, B., Sack, L. (2016). The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought. Proc. Natl. Acad. Sci. U.S.A. 113, 13098–13103. doi: 10.1073/pnas.1604088113
Bartlett, M. K., Scoffoni, C., Ardy, R., Zhang, Y., Sun, S., Cao, K., et al. (2012b). Rapid determination of comparative drought tolerance traits: Using an osmometer to predict turgor loss point. Methods Ecol. Evol. 3, 880–888. doi: 10.1111/j.2041-210X.2012.00230.x
Bartlett, M. K., Scoffoni, C., Sack, L. (2012a). The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol. Lett. 15, 393–405. doi: 10.1111/j.1461-0248.2012.01751.x
Bartlett, M. K., Sinclair, G. (2021a). Temperature and evaporative demand drive variation in stomatal and hydraulic traits across grape cultivars. J. Exp. Bot. 72, 1995–2009. doi: 10.1093/jxb/eraa577
Bartlett, M. K., Sinclair, G., Fontanesi, G., Knipfer, T., Walker, M. A., McElrone, A. J. (2021b). Root pressure-volume curve traits capture rootstock drought tolerance. Ann. Bot-London 129, 389–402. doi: 10.1093/aob/mcab132
Bauer, H., Ache, P., Lautner, S., Fromm, J., Hartung, W., Al-Rasheid, K. A., et al. (2013). The stomatal response to reduced relative humidity requires guard cellautonomous ABA synthesis. Curr. Biol. 23, 53–57. doi: 10.1016/j.cub.2012.11.022
Binks, O., Meir, P., Rowland, L., da Costa, A. C.L., Vasconcelos, S.S., de Oliveira, A. A.R., et al. (2016). Plasticity in leaf-level water relations of tropical rainforest trees in response to experimental drought. New Phytol. 211, 477–488. doi: 10.1111/nph.13927
Blackman, C. J., Li, X., Choat, B., Rymer, P. D., De Kauwe, M. G., Duursma, R. A., et al. (2019). Desiccation time during drought is highly predictable across species of eucalyptus from contrasting climates. New Phytol. 224, 632–643. doi: 10.1111/nph.16042
Bonan, G. B. (2008). Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science. 320, 1444–1449. doi: 10.1126/science.1155121
Bond, B. J., Kavanagh, K. L. (1999). Stomatal behavior of four woody species in relation to leaf-specific hydraulic conductance and threshold water potential. Tree Physiol. 19, 503—510. doi: 10.1093/treephys/19.8.503
Breshears, D. D., Myers, O. B., Meyer, C. W., Barnes, F. J., Zou, C. B.. (2009). Tree die-off in response to global change-type drought: Mortality insights from a decade of plant water potential measurements. Front. Ecol. Environ. 7, 185–189. doi: 10.1890/080016
Brodribb, T. J., Cochard, H. (2009). Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiol. 149, 575–584. doi: 10.1104/pp.108.129783
Brodribb, T. J., Holbrook, N. M. (2003). Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol. 132, 2166–2173. doi: 10.1104/pp.103.023879
Brodribb, T. J., McAdam, S. A. M. (2013). Abscisic acid mediates a divergence in the drought response of two conifers. Plant Physiol. 162, 1370–1377. doi: 10.1104/pp.113.217877
Brodribb, T. J., McAdam, S. A. M. (2015). Evolution in the smallest valves (stomata) guides even the biggest trees. Tree Physiol. 35, 451–452. doi: 10.1093/treephys/tpv042
Brodribb, T. J., McAdam, S. A. M., Jordan., G. J., Martins, S. C. V. (2014). Conifer species adapt to low-rainfall climates by following one of two divergent pathways. Proc. Natl. Acad. Sci. U.S.A. 111, 14489–14493. doi: 10.1073/pnas.1407930111
Brodribb, T. J., Powers, J., Cochard, H., Choat, B. (2020). Hanging by a thread? forests and drought. Science. 368, 261–266. doi: 10.1126/science.aat7631
Cai, G., Ahmed, M., Abdalla, M., Carminati, A. (2022). Root hydraulic phenotypes impacting water uptake in drying soils. Plant Cell Environ. 45, 650–663. doi: 10.1111/pce.14259
Cai, J., Tyree, M. T. (2010). The impact of vessel size on vulnerability curves: Data and models for within-species variability in saplings of aspen, Populus tremuloides michx. Plant Cell Environ. 33, 1059–1069. doi: 10.1111/j.1365-3040.2010.02127.x
Cardoso, A. A., Billon, L., Fanton Borges, A., Fernández-de-Uña, L., Gersony, J. T., Güney, A., et al. (2020). New developments in understanding plant water transport under drought stress. New Phytol. 227, 1025–1027. doi: 10.1111/nph.16663
Carmona, C. P., Bueno, C. G., Toussaint, A., Träger, S., Díaz, S., Moora, M., et al. (2021). Fine-root traits in the global spectrum of plant form and function. Nature 597, 683–687. doi: 10.1038/s41586-021-03871-y
Chen, Y., Choat, B., Sterck, F., Maenpuen, P., Katabuchi, M., Zhang, S., et al. (2021a). Hydraulic prediction of drought-induced plant dieback and top-kill depends on leaf habit and growth form. Ecol. Lett. 24, 2350–2363. doi: 10.1111/ele.13856
Chen, Z., Li, S., Luan, J., Zhang, Y., Zhu, S., Wan, X., et al. (2019a). Prediction of temperate broadleaf tree species mortality in arid limestone habitats with stomatal safety margins. Tree Physiol. 39, 1428–1437. doi: 10.1093/treephys/tpz045
Chen, Z., Liu, S., Lu, H., Wan, X. (2019b). Interaction of stomatal behaviour and vulnerability to xylem cavitation determines the drought response of three temperate tree species. AoB Plants 11, plz058. doi: 10.1093/aobpla/plz058
Chen, Z., Zhang, Y., Yuan, W., Zhu, S., Pan, R., Wan, X., et al. (2021b). Coordinated variation in stem and leaf functional traits of temperate broadleaf tree species in the isohydric-anisohydric spectrum. Tree Physiol. 41, 1601–1610. doi: 10.1093/treephys/tpab028
Chen, Z., Zhu, S., Zhang, Y., Luan, J., Li, S., Sun, P., et al. (2020). Tradeoff between storage capacity and embolism resistance in the xylem of temperate broadleaf tree species. Tree Physiol. 40, 1029–1042. doi: 10.1093/treephys/tpaa046
Chitra-Tarak, R., Xu, C., Aguilar, S., Anderson-Teixeira, K. J., Chambers, J., Detto, M., et al. (2021). Hydraulically-vulnerable trees survive on deep-water access during droughts in a tropical forest. New Phytol. 231, 1798–1813. doi: 10.1111/nph.17464
Choat, B., Brodribb, T. J., Brodersen, C. R., Duursma, R. A., López, R., Medlyn, B. E. (2018). Triggers of tree mortality under drought. Nature 558, 531–539. doi: 10.1038/s41586-018-0240-x
Choat, B., Cobb, A. R., Jansen, S. (2008). Structure and function of bordered pits: new discoveries and impacts on whole-plant hydraulic function. New Phytol. 177, 608–625. doi: 10.1111/j.1469-8137.2007.02317.x
Choat, B., Jansen, S., Brodribb, T. J. (2012). Global convergence in the vulnerability of forests to drought. Nature. 491, 752–756. doi: 10.1038/nature11688
Cochard, H., Badel, E., Herbette, S., Delzon, S., Choat, B., Jansen, S. (2013). Methods for measuring plant vulnerability to cavitation: a critical review. J. Exp. Bot. 64, 4779–4791. doi: 10.1093/jxb/ert193
Cowan, I. R., Farquhar, G. D. (1977). Stomatal function in relation to leaf metabolism and environment. Symp. Soc. Exp. Biol. 31, 475–505.
Cuneo, I. F., Knipfer, T., Brodersen, C. R., McElrone, A. J. (2016). Mechanical failure of fine root cortical cells initiates plant hydraulic decline during drought. Plant Physiol. 172, 1669–1678. doi: 10.1104/pp.16.00923
Cuneo, I. F., Knipfer, T., Mandal, P., Brodersen, C. R., McElrone, A. J. (2018). Water uptake can occur through woody portions of roots and facilitates localized embolism repair in grapevine. New Phytol. 218, 506–516. doi: 10.1111/nph.15032
Davies, W. J., Zhang, J. (1991). Root signals and the regulation of growth and development of plants in drying soil. Annu. Rev. Plant Physiol. Mol. Biol. 42, 55–76. doi: 10.1146/annurev.pp.42.060191.000415
Dixon, H. H., Joly, J. (1895). On the ascent of sap. Philosophical Transactions of the Royal Society of London B, 186, 563–576.
Dodd, I. C., Egea, G., Watts, C. W., Whalley, W. R. (2010). Root water potential integrates discrete soil physical properties to influence ABA signalling during partial rootzone drying. J. ExpBot. 61, 3543–3551. doi: 10.1093/jxb/erq195
Drake, P. L., Price, C. A., Poot, P., Veneklaas, E. J. (2015). Isometric partitioning of hydraulic conductance between leaves and stems: balancing safety and efficiency in different growth forms and habitats. Plant Cell Environ. 38, 1628–1636. doi: 10.1111/pce.12511
Edwards, D., Kerp, H., Hass, H. (1998). Stomata in early land plants: An anatomical and ecophysiological approach. J. Exp. Bot. 49, 255–278. doi: 10.1093/jxb/49.Special_Issue.255
Eller, C. B., Lima, A. L., Oliveira, R. S. (2013). Foliar uptake of fog water and transport belowground alleviates drought effects in the cloud forest tree species, drimys brasiliensis (Winteraceae). New Phytol. 199, 151–162. doi: 10.1111/nph.12248
Fichot, R., Brignolas, F., Cochard, H., Ceulemans, R. (2015). Vulnerability to drought-induced cavitation in poplars: synthesis and future opportunities. Plant Cell Environ. 38, 1233–1251. doi: 10.1111/pce.12491
Fu, X., Meinzer, F. C. (2019). Metrics and proxies for stringency of regulation of plant water status (iso/anisohydry): A global data set reveals coordination and tradeoffs among water transport traits. Tree Physiol. 39, 122–134. doi: 10.1093/treephys/tpy087
Fu, X., Meinzer, F. C., Woodruff, D. R., Liu, Y., Smith, D. D., McCulloh, K. A., et al. (2019). Coordination and trade-offs between leaf and stem hydraulic traits and stomatal regulation along a spectrum of isohydry to anisohydry. Plant Cell Environ. 42, 2245–2258. doi: 10.1111/pce.13543
Garzón, M. B., González Muñoz, N., Wigneron, J., Moisy, C., Fernández-Manjarrés, J., Delzon, S. (2018). The legacy of water deficit on populations having experienced negative hydraulic safety margin. Global Ecol. Biogeogr. 27, 346–356. doi: 10.1111/geb.12701
Hacke, U. G., Sperry, J. S. (2001). Functional and ecological xylem anatomy. Perspect. Plant Ecol. 4, 97–115. doi: 10.1078/1433-8319-00017
Hacke, U. G., Sperry, J. S., Wheeler, J. K., Castro, L. (2006). Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol. 26, 689–701. doi: 10.1093/treephys/26.6.689
Hacke, U. G., Stiller, V., Sperry, J. S., Pittermann, J., McCulloh, K. A. (2001). Cavitation fatigue. embolism and refilling cycles can weaken the cavitation resistance of xylem. Plant Physiol. 125, 779–786. doi: 10.1104/pp.125.2.779
Hafner, B. D., Hesse, B. D., Bauerle, T. L., Grams, T. E. E. (2020). Water potential gradient, root conduit size and root xylem hydraulic conductivity determine the extent of hydraulic redistribution in temperate trees. Funct. Ecol. 34, 561–574. doi: 10.1111/1365-2435.13508
Hammond, W. M., Adams, H. D. (2019). Dying on time: Traits influencing the dynamics of tree mortality risk from drought. Tree Physiol. 39, 906–909. doi: 10.1093/treephys/tpz050
Hartmann, H., Moura, C. F., Anderegg, W. R. L., Ruehr, N. K., Salmon, Y., Allen, C. D., et al. (2018). Research frontiers for improving our understanding of drought-induced tree and forest mortality. New Phytol. 218, 15–28. doi: 10.1111/nph.15048
Hartmann, H., Roman, M. L., Schuldt, B. (2021). A whole-plant perspective of isohydry: stem-level support for leaf-level plant water regulation. Tree Physiol. 41, 901–905. doi: 10.1093/treephys/tpab011
Harvey, H. P., Van Den Driessche, R. (1997). Nutrition, xylem cavitation and drought resistance in hybrid poplar. Tree Physiol. 17, 647–654. doi: 10.1093/treephys/17.10.647
Heath, O. V. S. (1938). An experimental investigation of the mechanism of stomatal movement, with some preliminary observations upon the response of the guard cells to “shock.” New Phytol. 37, 385–395. doi: 10.1111/j.1469-8137.1938.tb06954.x
Hedwig, J. (1793). Sammlung seiner zerstreuten abhandlungen und beobachtungen über botanisch-ökonomische gegenstände (Leipzig, Germany: Band 1. Siegfried Lebrecht Crucius).
Henry, C., John, G. P., Pan, R., Bartlett, M. K., Fletcher, L. R., Scoffoni, C., et al. (2019). A stomatal safety-efficiency trade-off constrains responses to leaf dehydration. Nat. Commun. 10, 1–9. doi: 10.1038/s41467-019-11006-1
Hevia, A., Sánchez-Salguero, R., Julio Camarero, J., Querejeta, J. I., Sangüesa-Barreda, G, Gazol, A. (2019). Long-term nutrient imbalances linked to drought-triggered forest dieback. Sci. Total Environ. 690, 1254–1267. doi: 10.1016/j.scitotenv.2019.06.515
Hochberg, U., Rockwell, F. E., Holbrook, N. M., Cochard, H. (2018). Iso/Anisohydry: A plant–environment interaction rather than a simple hydraulic trait. Trends Plant Sci. 23, 112–120. doi: 10.1016/j.tplants.2017.11.002
Ishida, A., Diloksumpun, S., Ladpala, P., Staporn, D., Panuthai, S., Gamo, M., et al. (2006). Contrasting seasonal leaf habits of canopy trees between tropical dry-deciduous and evergreen forests in Thailand. Tree Physiol. 26, 643–656. doi: 10.1093/treephys/26.5.643
Jacobsen, A. L., Agenbag, L., Esler, K. J., Pratt, R. B., Ewers, F. W., Davis, S. D. (2007). Xylem density, biomechanics and anatomical traits correlate with water stress in 17 evergreen shrub species of the Mediterranean-type climate region of south. Africa. J. Ecol. 95, 171–183. doi: 10.1111/j.1365-2745.2006.01186.x
Jacobsen, A. L., Ewers, F. W., Pratt, R. B., Paddock, W. A., Davis, S. D. (2005). Do xylem fibers affect vessel cavitation resistance? Plant Physiol. 139, 546–556. doi: 10.1104/pp.104.058404
Johnson, D. M., Berry, Z. C., Baker, K. V., Smith, D. D., McCulloh, K. A., Domec, J. (2018). Leaf hydraulic parameters are more plastic in species that experience a wider range of leaf water potentials. Funct. Ecol. 32, 894–903. doi: 10.1111/1365-2435.13049
Johnson, D. M., McCulloh, K. A., Woodruff, D. R., Meinzer, F. C. (2012). Hydraulic safety margins and embolism reversal in stems and leaves: why are conifers and angiosperms so different? Plant Sci. 195, 48–53. doi: 10.1016/j.plantsci.2012.06.010
Kaack, L., Weber, M., Isasa, E., Karimi, Z., Li, S., Pereira, L., et al. (2021). Pore constrictions in intervessel pit membranes provide a mechanistic explanation for xylem embolism resistance in angiosperms. New Phytol. 230, 1829–1843. doi: 10.1111/nph.17282
Klein, T. (2014). The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Funct. Ecol. 28, 1313–1320. doi: 10.1111/1365-2435.12289
Koebernick, N., Huber, K., Kerkhofs, E., Vanderborght, J., Mathieu, J., Vereecken, H., et al. (2015). Unraveling the hydrodynamics of split root water uptake experiments using CT scanned root architectures and three dimensional flow simulations. Front. Plant Sci. 6, 370. doi: 10.3389/fpls.2015.00370
Kollist, H., Nuhkat, M., Roelfsema, M. R. G. (2014). Closing gaps: Linking elements that control stomatal movement. New Phytol. 203, 44–62. doi: 10.1111/nph.12832
Kozlowski, T. T., Pallardy, S. G. (2002). Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 68, 270–334. doi: 10.1663/0006-8101(2002)068[0270:AAAROW]2.0.CO;2
Kuromori, T., Sugimoto, E., Shinozaki, K. (2014). Intertissue signal transfer of abscisic acid from vascular cells to guard cells. Plant Physiol. 164, 1587–1592. doi: 10.1104/pp.114.235556
Lamy, J. B., Delzon, S., Bouche, P. S., Alia, R., Giuseppe Vendramin, G., Cochard, H., et al. (2014). Limited genetic variability and phenotypic plasticity detected for cavitation resistance in a Mediterranean pine. New Phytol. 201, 874–886. doi: 10.1111/nph.12556
Larter, M., Brodribb, T. J., Pfautsch, S., Burlett, R., Cochard, H., Delzon, S. (2015). Extreme aridity pushes trees to their physical limits. Plant Physiol. 168, 804–807. doi: 10.1104/pp.15.00223
Larter, M., Pfautsch, S., Domec, J., Trueba, S., Nagalingum, N., Delzon, S. (2017). Aridity drove the evolution of extreme embolism resistance and the radiation of conifer genus callitris. New Phytol. 215, 97–112. doi: 10.1111/nph.14545
Lawson, T., Blatt, M. R. (2014). Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency. Plant Physiol. 164, 1556–1570. doi: 10.1104/pp.114.237107
Lee, K. H., Piao, H. L., Kim, H. Y., Choi, S. M., Jiang, F., Hartung, W., et al. (2006). Activation of glucosidase via stressinduced polymerization rapidly increases active pools of abscisic acid. Cell. 126, 1109–1120. doi: 10.1016/j.cell.2006.07.034
Lenz, T. I., Wright, I. J., Westoby, M. (2006). Interrelations among pressure-volume curve traits across species and water availability gradients. Physiol. Plant 127, 423–433. doi: 10.1111/j.1399-3054.2006.00680.x
Li, X., Blackman, C. J., Peters, J. M. R., Choat, B., Rymer, P. D., Medlyn, B. E., et al. (2019). More than iso/anisohydry: Hydroscapes integrate plant water use and drought tolerance traits in 10 eucalypt species from contrasting climates. Funct. Ecol. 33, 1035–1049. doi: 10.1111/1365-2435.13320
Li, S., Feifel, M., Karimi, Z., Schuldt, B., Choat, B., Jansen, S. (2015). Leaf gas exchange performance and the lethal water potential of five European species during drought. Tree Physiol. 36, 179–192. doi: 10.1093/treephys/tpv117
Li, S., Lens, F., Espino, S., Karimi, Z., Klepsch, M. M., Schenk, H. J., et al. (2016). Intervessel pit membrane thickness as a key determinant of embolism resistance in angiosperm xylem. IAWA J. 37, 152–171. doi: 10.1163/22941932-20160128
Li, S., Li, X., Wang, J., Chen, Z., Lu, S., Wan, X., et al. (2022). Hydraulic traits are coupled with plant anatomical traits under drought-rewatering cycles in Ginkgo biloba l. Tree Physiol. 42, 1216–1227. doi: 10.1093/treephys/tpab174
Limousin, J. M., Longepierre, D., Huc, R., Rambal, S. (2010). Change in hydraulic traits of Mediterranean quercus ilex subjected to long-term throughfall exclusion. Tree Physiol. 30, 1026–1036. doi: 10.1093/treephys/tpq062
Lynch, J. P. (2019). Root phenotypes for improved nutrient capture: An underexploited opportunity for global agriculture. New Phytol. 223, 548–564. doi: 10.1111/nph.15738
Maherali, H., Pockman, W., Jackson, R. (2004). Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology. 85, 2184–2199. doi: 10.1890/02-0538
Maréchaux, I., Bartlett, M. K., Iribar, A., Sack, L., Chave, J. (2017). Stronger seasonal adjustment in leaf turgor loss point in lianas than trees in an Amazonian forest. Biol. Lett. 13, 20160819. doi: 10.1098/rsbl.2016.0819
Markesteijn, L., Poorter, L. (2011). Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traitspc. Plant Cell Environ. 34, 137–148. doi: 10.1111/j.1365-3040.2010.02231.x
Martínez-Vilalta, J., Garcia-Forner, N. (2017). Water potential regulation, stomatal behaviour and hydraulic transport under drought: Deconstructing the iso/anisohydric concept. Plant Cell Environ. 40, 962–976. doi: 10.1111/pce.12846
Martínez-Vilalta, J., Rafael, P., Aguadé, D., Javier, R., Mencuccini, M. (2014). A new look at water transport regulation in plants. New Phytol. 204, 105–115. doi: 10.1111/nph.12912
Martin-StPaul, N., Delzon, S., Cochard, H. (2017). Plant resistance to drought depends on timely stomatal closure. Ecol. Lett. 20, 1437–1447. doi: 10.1111/ele.12851
McAdam, S. A. M., Brodribb, T. J. (2015). The evolution of mechanisms driving the stomatal response to vapor pressure deficit. Plant Physiol. 167, 833–843. doi: 10.1104/pp.114.252940
McCormack, M. L., Dickie, I. A., Eissenstat, D. M., Fahey, T. J., Fernandez, C. W., Guo, D., et al. (2015). Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 207, 505–518. doi: 10.1111/nph.13363
Mccullon, K. A., Johnson, D. M., Meinzer, F. C. (2012). Hydraulic architecture of two species differing in wood density: Opposing strategies in co-occurring tropical pioneer trees. Plant Cell Environ. 35, 116–125. doi: 10.1111/j.1365-3040.2011.02421.x
McDowell, N. G. (2011). Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 155, 1051–1059. doi: 10.1104/pp.110.170704
McDowell, N., Pockman, W. T., Allen, C. D., Breshears, D. D., Cobb, N., Kolb, T., et al. (2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 178, 719–739. doi: 10.1111/j.1469-8137.2008.02436.x
McKown, A. D., Cochard, H., Sack, L. (2010). Decoding leaf hydraulics with a spatially explicit model: principles of venation architecture and implications for its evolution. Am. Nat. 175, 447–460. doi: 10.1086/650721
Medeiros, D. B., Daloso, D. M., Fernie, A. R., Nikoloski, Z., Araújo, W. L. (2015). Utilizing systems biology to unravel stomatal function and the hierarchies underpinning its control. Plant Cell Environ. 38, 1457–1470. doi: 10.1111/pce.12517
Meinzer, F. C., Johnson, D. M., Lachenbruch, B., McCulloh, K. A., Woodruff, D. R. (2009). Xylem hydraulic safety margins in woody plants: coordination of stomatal control of xylem tension with hydraulic capacitance. Funct. Ecol. 23, 922–930. doi: 10.1111/j.1365-2435.2009.01577.x
Meinzer, F. C., McCulloh, K. A. (2013). Xylem recovery from drought-induced embolism: where is the hydraulic point of no return? Tree Physiol. 33, 331–334. doi: 10.1093/treephys/tpt022
Meinzer, F. C., Sharifi, M. R., Nilsen, E. T., Rundel, P. W. (1988). Effects of manipulation of water and nitrogen regime on the water relations of the desert shrub Larrea tridentata. Oecologia. 77, 480–486. doi: 10.1007/BF00377263
Meinzer, F. C., Smith, D. D., Woodruff, D. R., Marias, D. E., McCulloh, K. A., Howard, A. R., et al. (2017). Stomatal kinetics and photosynthetic gas exchange along a continuum of isohydric to anisohydric regulation of plant water status. Plant Cell Environ. 40, 1618–1628. doi: 10.1111/pce.12970
Meinzer, F. C., Woodruff, D. R., Marias, D. E., McCulloh, K. A., Sevanto, S. (2014). Dynamics of leaf water relations components in co-occurring iso-and anisohydric conifer species. Plant Cell Environ. 37, 2577–2586. doi: 10.1111/pce.12327
Meinzer, F. C., Woodruff, D. R., Marias, D. E., Smith, D. D., McCulloh, K. A., Howard, A. R., et al. (2016). Mapping ‘hydroscapes’ along the iso- to anisohydric continuum of stomatal regulation of plant water status. Ecol. Lett. 19, 1343–1352. doi: 10.1111/ele.12670
Nardini, A., Luglio, J. (2014). Leaf hydraulic capacity and drought vulnerability: Possible trade-offs and correlations with climate across three major biomes. Funct. Ecol. 28, 810–818. doi: 10.1111/1365-2435.12246
Nelson, S. K., Oliver, M. J. (2017). A soil-plate based pipeline for assessing cereal root growth in response to polyethylene glycol (PEG)-induced water deficit stress. Front. Plant Sci. 8, 1272. doi: 10.3389/fpls.2017.01272
Nolan, R. H., Tarin, T., Santini, N. S., McAdam, S. A. M., Ruman, R., Eamus, D. (2017). Differences in osmotic adjustment, foliar abscisic acid dynamics and stomatal regulation between an isohydric and anisohydric woody angiosperm during drought. Plant Cell Environ. 40, 3122–3134. doi: 10.1111/pce.13077
Nolf, M., Creek, D., Duursma, R., Holtum, J., Mayr, S., Choat, B. (2015). Stem and leaf hydraulic properties are finely coordinated in three tropical rainforest tree species. Plant Cell Environ. 38, 2652–2661. doi: 10.1111/pce.12581
Okamoto, M., Tanaka, Y., Abrams, S. R., Kamiya, Y., Seki, M., Nambara, E. (2009). High humidity induces abscisic acid 89-hydroxylase in stomata and vasculature to regulate local and systemic abscisic acid responses in arabidopsis. Plant Physiol. 149, 825–834. doi: 10.1104/pp.108.130823
Oliveira, R. S., Eller, C. B., Barros, F. V., Hirota, M., Brum, M., Bittencourt, P. (2021). Linking plant hydraulics and the fast-slow continuum to understand resilience to drought in tropical ecosystems. New Phytol. 230, 904–923. doi: 10.1111/nph.17266
Pan, Y., Birdsey, R. A., Fang, J., Houghton, R., Kauppi, P. E., Kurz, W. A., et al. (2011). A large and persistent carbon sink in the world’s forests. Science. 333, 988–993. doi: 10.1126/science.1201609
Papanatsiou, M., Petersen, J., Henderson, L., Wang, Y., Christie, J. M., Blatt, M. R. (2019). Optogenetic manipulation of stomatal kinetics improves carbon assimilation, water use, and growth. Science. 363, 1456–1459. doi: 10.1126/science.aaw0046
Phillips, O. L., Aragão, L. E. O. C., Lewis, S. L., Fisher, J. B., Lloyd, J., López-González, G., et al. (2009). Drought sensitivity of the amazon rainforest. Science. 323, 1344–1347. doi: 10.1126/science.1164033
Pivovaroff, A. L., Cook, V. M. W., Santiago, L. S. (2018). Stomatal behaviour and stem xylem traits are coordinated for woody plant species under exceptional drought conditions. Plant Cell Environ. 41, 2617–2626. doi: 10.1111/pce.13367
Pivovaroff, A. L., Pasquini, S. C., De Guzman, M. E., Alstad, K. P., Stemke, J. S., Santiago, L. S. (2016). Multiple strategies for drought survival among woody plant species. Funct. Ecol. 30, 517–526. doi: 10.1111/1365-2435.12518
Plavcová, L., Hacke, U. G. (2012). Phenotypic and developmental plasticity of xylem in hybrid poplar saplings subjected to experimental drought, nitrogen fertilization, and shading. J. Exp. Bot. 63, 6481–6491. doi: 10.1093/jxb/ers303
Plavcová, L., Hacke, U. G., Almeida-Rodriguez, A. M., Li, E., Douglas, C. J. (2013). Gene expression patterns underlying changes in xylem structure and function in response to increased nitrogen availability in hybrid poplar. Plant Cell Environ. 36, 186–199. doi: 10.1111/j.1365-3040.2012.02566.x
Plavcová, L., Hacke, U. G., Sperry, J. S. (2011). Linking irradiance-induced changes in pit membrane ultrastructure with xylem vulnerability to cavitation. Plant Cell Environ. 34, 501–513. doi: 10.1111/j.1365-3040.2010.02258.x
Raven, J. A. (2002). Selection pressures on stomatal evolution. New Phytol. 153, 371–386. doi: 10.1046/j.0028-646X.2001.00334.x
Rowland, L., Martínez-Vilalta, J., Mencuccini, M. (2021). Hard times for high expectations from hydraulics: Predicting drought-induced forest mortality at landscape scales remains a challenge. New Phytol. 230, 1685–1687. doi: 10.1111/nph.17317
Sack, L., Buckley, T. N. (2016). The developmental basis of stomatal density and flux. Plant Physiol. 171, 2358–2363. doi: 10.1104/pp.16.00476
Salleo, S., Nardini, A., Pitt, F., Lo Gullo, M. A. (2000). Xylem cavitation and hydraulic control of stomatal conductance in laurel (Laurus nobilis l.). Plant Cell Environ. 23, 71–79. doi: 10.1046/j.1365-3040.2000.00516.x
Savi, T., Bertuzzi, S., Branca, S., Tretiach, M., Nardini, A. (2015). Drought-induced xylem cavitation and hydraulic deterioration: Risk factors for urban trees under climate change? New Phytol. 205, 1106–1116. doi: 10.1111/nph.13112
Scholander, P. F., Bradstreet, E. D., Hemmingsen, E. A., Hammel, H. T. (1965). Sap pressure in vascular plants. Science. 148, 339–346. doi: 10.1126/science.148.3668.339
Scholz, A., Rabaey, D., Stein, A., Cochard, H., Smets, E., Jansen, S. (2013). The evolution and function of vessel and pit characters with respect to cavitation resistance across 10 prunus species. Tree Physiol. 33, 684–694. doi: 10.1093/treephys/tpt050
Schoonmaker, A. L., Hacke, U. G., Landhäusser, S. M., Lieffers, V. J., Tyree, M. T. (2010). Hydraulic acclimation to shading in boreal conifers of varying shade tolerance. Plant Cell Environ. 33, 382–393. doi: 10.1111/j.1365-3040.2009.02088.x
Schuldt, B., Knutzen, F., Delzon, S., Jansen, S., Muller-Haubold, H., Burlett, R., et al. (2016). How adaptable is the hydraulic system of European beech in the face of climate change-related precipitation reduction? New Phytol. 210, 443–458. doi: 10.1111/nph.13798
Scoffoni, C., Vuong, C., Diep, S., Cochard, H., Sack, L. (2014). Leaf shrinkage with dehydration: coordination with hydraulic vulnerability and drought tolerance. Plant Physiol. 164, 1772–1788. doi: 10.1104/pp.113.221424
Shabala, S. N., Lew, R. R. (2002). Turgor regulation in osmotically stressed arabidopsis epidermal root cells. direct support for the role of inorganic ion uptake as revealed by concurrent flux and cell turgor measurements. Plant Physiol. 129, 290–299. doi: 10.1104/pp.020005
Skelton, R. P., Westa, A. G., Dawson, T. E. (2015). Predicting plant vulnerability to drought in biodiverse regions using functional traits. Proc. Natl. Acad. Sci. U.S.A. 112, 5744–5749. doi: 10.1073/pnas.1503376112
Slatyer, R. O., Taylor, S. A. (1960). Terminology in plant- and soil-water relations. Nature. 187, 922–924. doi: 10.1038/187922a0
Sperry, J. S., Perry, A. H., Sullivan, J. E. M. (1991). Pit membrane degradation and air-embolism formation in ageing xylem vessels of Populus tremuloides michx. J. Exp. Bot. 42, 1399–1406. doi: 10.1093/jxb/42.11.1399
Tardieu, F. (2016). Too many partners in root-shoot signals. does hydraulics qualify as the only signal that feeds back over time for reliable stomatal control? New Phytol. 212, 802–804. doi: 10.1111/nph.14292
Tardieu, F., Draye, X., Javaux, M. (2017). Root water uptake and ideotypes of the root system: Whole-plant controls matter. Vadose Zone J. 16, 9. doi: 10.2136/vzj2017.05.0107
Tardieu, F., Simonneau, T. (1998). Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours. J. Exp. Bot. 49, 419–432. doi: 10.1093/jxb/49.Special_Issue.419
Torres-Ruiz, J. M., Cochard, H., Fonseca, E., Badel, E., Gazarini, L., Vaz, M. (2017). Differences in functional and xylem anatomical features allow cistus species to co-occur and cope differently with drought in the Mediterranean region. Tree Physiol. 37, 755–766. doi: 10.1093/treephys/tpx013
Tracy, S. R., Nagel, K. A., Postma, J. A., Fassbender, H., Wasson, A., Watt, M. (2020). Crop improvement from phenotyping roots: Highlights reveal expanding opportunities. Trends Plant Sci. 25, 105–118. doi: 10.1016/j.tplants.2019.10.015
Trugman, A. T., Anderegg, L. D. L., Anderegg, W. R. L., Das, A. J., Stephenson, N. L. (2021). Why is tree drought mortality so hard to predict? Trends Ecol. Evol. 36, 520–532. doi: 10.1016/j.tree.2021.02.001
Tyree, M. T., Ewers, F. W. (1991). The hydraulic architecture of trees and other woody plants. New Phytol. 119, 345–360. doi: 10.1111/j.1469-8137.1991.tb00035.x
Tyree, M. T., Sperry, J. S. (1988). Do woody plants operate near the point of catastrophic xylem dysfunction caused by dynamic water stress? Plant Physiol. 88, 574–580. doi: 10.1104/pp.88.3.574
Tyree, M. T., Zimmermann, M. H. (2002). Xylem structure and the ascent of sap, 2nd edn (Berlin, Germany: Springer).
Urli, M., Porté, A. J., Cochard, H., Guengant, Y., Burlett, R., Delzon, S. (2013). Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree Physiol. 33, 672–683. doi: 10.1093/treephys/tpt030
Waadt, R., Hitomi, K., Nishimura, N., Hitomi, C., Adams, S. R., Getzoff, E. D., et al. (2014). FRET-based reporters for the direct visualization of abscisic acid concentration changes and distribution in arabidopsis. eLife. 3, e01739. doi: 10.7554/eLife.01739
Wasaya, A., Zhang, X., Fang, Q., Yan, Z. (2018). Root phenotyping for drought tolerance: A review. Agronomy 8, 241. doi: 10.3390/agronomy8110241
Wheeler, J. K., Sperry, J. S., Hacke, U. G., Hoang, N. (2005). Inter-vessel pitting and cavitation in woody rosaceae and other vesselled plants: A basis for a safety versus efficiency trade-off in xylem transport. Plant Cell Environ. 28, 800–812. doi: 10.1111/j.1365-3040.2005.01330.x
Woodruff, D. R., Meinzer, F. C., Marias, D. E., Sevanto, S., Jenkins, M. W., McDowell, N. G. (2015). Linking nonstructural carbohydrate dynamics to gas exchange and leaf hydraulic behavior in pinus edulis and juniperus monosperma. New Phytol. 206, 411–421. doi: 10.1111/nph.13170
Wortemann, R., Herbette, S., Barigah, T. S., Fumanal, B., Alia, R., Ducousso, A., et al. (2011). Genotypic variability and phenotypic plasticity of cavitation resistance in fagus sylvatica l. across Europe. Tree Physiol. 31, 1175–1182. doi: 10.1093/treephys/tpr101
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). The worldwide leaf economics spectrum. Nature. 428, 821–827. doi: 10.1038/nature02403
Wu, T., Tissue, D. T., Li, X., Liu, S., Chu, G., Zhou, G., et al. (2020). Long-term effects of 7-year warming experiment in the field on leaf hydraulic and economic traits of subtropical tree species. Glob. Change Biol. 26, 7144–7157. doi: 10.1111/gcb.15355
Yamauchi, T., Pedersen, O., Nakazono, M., Tsutsumi, N. (2020). Key root traits of poaceae for adaptation to soil water gradients. New Phytol. 229, 3133–3140. doi: 10.1111/nph.17093
Keywords: climate change, hydraulic traits, hydraulic failure, carbon starvation, stomatal safety margin, tree mortality
Citation: Chen Z, Li S, Wan X and Liu S (2022) Strategies of tree species to adapt to drought from leaf stomatal regulation and stem embolism resistance to root properties. Front. Plant Sci. 13:926535. doi: 10.3389/fpls.2022.926535
Received: 22 April 2022; Accepted: 12 September 2022;
Published: 27 September 2022.
Edited by:
Teemu Hölttä, University of Helsinki, FinlandReviewed by:
Edgard Picoli, Universidade Federal de Viçosa, BrazilLouis S. Santiago, University of California, Riverside, United States
Copyright © 2022 Chen, Li, Wan and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shirong Liu, liusr@caf.ac.cn