

Abstract 摘要
Subarachnoid hemorrhage (SAH) is a devastating subtype of stroke with high mortality and disability rate. Meningeal lymphatic vessels (mLVs) are a newly discovered intracranial fluid transport system and are proven to drain extravasated erythrocytes from cerebrospinal fluid into deep cervical lymph nodes after SAH. However, many studies have reported that the structure and function of mLVs are injured in several central nervous system diseases. Whether SAH can cause mLVs injury and the underlying mechanism remain unclear. Herein, single‐cell RNA sequencing and spatial transcriptomics are applied, along with in vivo/vitro experiments, to investigate the alteration of the cellular, molecular, and spatial pattern of mLVs after SAH. First, it is demonstrated that SAH induces mLVs impairment. Then, through bioinformatic analysis of sequencing data, it is discovered that thrombospondin 1 (THBS1) and S100A6 are strongly associated with SAH outcome. Furthermore, the THBS1‐CD47 ligand‐receptor pair is found to function as a key role in meningeal lymphatic endothelial cell apoptosis via regulating STAT3/Bcl‐2 signaling. The results illustrate a landscape of injured mLVs after SAH for the first time and provide a potential therapeutic strategy for SAH based on mLVs protection by disrupting THBS1 and CD47 interaction.
蛛网膜下腔出血(SAH)是一种破坏性极强的中风亚型,具有高死亡率和高致残率。脑膜淋巴管(mLVs)是新近发现的颅内液体运输系统,已被证实能在 SAH 后将渗出的红细胞从脑脊液引流至深部颈淋巴结。然而,多项研究报道,mLVs 的结构和功能在多种中枢神经系统疾病中受损。SAH 是否会导致 mLVs 损伤及其潜在机制尚不明确。本研究结合单细胞 RNA 测序与空间转录组学技术,并通过体内外实验,探究 SAH 后 mLVs 在细胞、分子及空间模式上的变化。首先证实 SAH 会导致 mLVs 功能损伤,随后通过测序数据的生物信息学分析发现,血小板反应蛋白 1(THBS1)和 S100A6 与 SAH 预后密切相关。进一步研究发现,THBS1-CD47 配体-受体对通过调控 STAT3/Bcl-2 信号通路,在脑膜淋巴管内皮细胞凋亡中发挥关键作用。 研究结果首次揭示了蛛网膜下腔出血(SAH)后脑膜淋巴管(mLVs)损伤的全貌,并提出了一种基于通过阻断 THBS1 与 CD47 相互作用保护 mLVs 的潜在 SAH 治疗策略。
Keywords: meningeal lymphatic, meningeal lymphatic endothelial cells, single‐cell RNA sequencing, spatial transcriptome, subarachnoid hemorrhage
关键词:脑膜淋巴管,脑膜淋巴管内皮细胞,单细胞 RNA 测序,空间转录组,蛛网膜下腔出血
This study integrates single‐cell RNA sequencing and spatial transcriptomics to comprehensively characterize the composition and alterations of different cell clusters in meningeal lymphatic vessels (mLVs) tissues after subarachnoid hemorrhage (SAH) in mice models for the first time. Thrombospondin 1‐CD47 ligand‐receptor interaction is proven to induce meningeal lymphatic endothelial cell apoptosis, which might be the initial reason for mLVs injury after SAH.
本研究首次整合单细胞 RNA 测序与空间转录组学技术,全面表征了小鼠模型中蛛网膜下腔出血(SAH)后脑膜淋巴管(mLVs)组织内不同细胞簇的组成与变化。研究证实,血小板反应蛋白 1-CD47 配体-受体相互作用可诱导脑膜淋巴管内皮细胞凋亡,这可能是 SAH 后 mLVs 损伤的初始原因。
1. Introduction 1. 引言
Subarachnoid hemorrhage (SAH) is a devastating subtype of stroke, characterized by a high rate of disability and mortality.[
1
] Early brain injury (EBI) and delayed cerebral ischemia (DCI) are the two major pathological processes after SAH. Regrettably, current therapies targeting both EBI and DCI show limited clinical benefits.[
2
] Hence, there is an urgent need to develop novel therapeutic strategies to promote the neurological function recovery of SAH patients. Evidence has indicated that the extravasation of erythrocytes in cerebrospinal fluid (CSF) is crucial to the SAH progress, which severely disturbs the CSF circulation in both acute and late phases after SAH. CSF circulation dysfunction can directly lead to brain edema formation and hydrocephalus through increasing intracerebral pressure (ICP) and decreasing cerebral blood perfusion.[
3
] Considering these SAH pathologies, accelerating extravasated blood clearance in subarachnoid space (SAS) may be an effective therapeutic approach.
蛛网膜下腔出血(SAH)是一种破坏性的中风亚型,具有高致残率和死亡率的特点。 [
1
] 早期脑损伤(EBI)和迟发性脑缺血(DCI)是 SAH 后两大主要病理过程。遗憾的是,目前针对 EBI 和 DCI 的治疗方案临床获益有限。 [
2
] 因此,迫切需要开发新的治疗策略以促进 SAH 患者神经功能恢复。有证据表明,脑脊液(CSF)中红细胞的渗出对 SAH 进展至关重要,其在 SAH 后的急性期和晚期均严重干扰了 CSF 循环。CSF 循环功能障碍可通过增加颅内压(ICP)和降低脑血流灌注,直接导致脑水肿形成和脑积水。 [
3
] 鉴于这些 SAH 病理变化,加速蛛网膜下腔(SAS)中渗出血液的清除可能是一种有效的治疗方法。
Lymphatic drainage plays an important role in modulating tissue homeostasis, excess fluid clearance and macromolecules, and immune cell migration.[
4
] The meningeal lymphatic system within the dura mater transports macromolecules away from the brain parenchyma and delivers CSF to the surrounding cervical lymph nodes.[
5
] Weakening of this central nervous system (CNS) drainage system leads to impairment of pathological substance clearance and is associated with neurodegenerative and age‐associated neurological conditions.[
6
] Subsequent studies have revealed that meningeal lymphatic vessels (mLVs) are responsible for the clearance of β‐amyloid and α‐synuclein, which lead to neurological deficits in Alzheimer's and Parkinson's diseases.[
7
] Recently, mLVs have also been reported to drain extravasated erythrocytes from CSF into deep cervical lymph nodes (dCLNs) after SAH.[
3b
] These findings further demonstrate that mLVs play key roles in the clearance of intracranial waste. However, little is known about the cellular architecture and underlying gene regulatory features of mLVs, especially after SAH.
淋巴引流在调节组织稳态、清除多余液体和大分子以及免疫细胞迁移中扮演重要角色。 [
4
] 硬脑膜内的脑膜淋巴系统负责将大分子从脑实质中运出,并将脑脊液输送至周围颈淋巴结。 [
5
] 中枢神经系统(CNS)这一引流系统的弱化会导致病理物质清除功能受损,并与神经退行性疾病及年龄相关神经病变相关联。 [
6
] 后续研究揭示,脑膜淋巴管(mLVs)负责清除β-淀粉样蛋白和α-突触核蛋白,这些蛋白的积累会导致阿尔茨海默病和帕金森病的神经功能缺损。 [
7
] 近期还有报道指出,SAH 后,mLVs 能将外渗的红细胞从脑脊液引流至深颈淋巴结(dCLNs)。 [
3b
] 这些发现进一步证实了 mLVs 在清除颅内废物中的关键作用。然而,关于 mLVs 的细胞结构及其基因调控特征,尤其是在 SAH 后的变化,目前知之甚少。
Single‐cell RNA sequencing (scRNA‐Seq) makes it possible to identify cell subpopulations and define their unique function by exploring the unique transcriptomic profile of each single cell, which therefore has emerged as a powerful tool to further explain pathophysiological changes in diseases.[
8
] However, information on spatial distribution and correlation with other cells of one individual cell is lost during tissue dissociation. Spatial transcriptomics (ST) is currently invented to identify cell types and cell‐to‐cell interactions on a structural and spatial level in a selected tissue, which can compensate for the drawbacks of scRNA‐seq.[
9
] Integration of scRNA‐Seq with ST enables in situ reproduction and visualization of pathologies in injured tissue than scRNA‐Seq or ST alone,[
10
] and has been applied in various tumor diseases.[
11
] Therefore, by integrating these two techniques, we will be able to better understand mLVs pathophysiology after SAH.
单细胞 RNA 测序(scRNA‐Seq)通过探索每个单细胞独特的转录组图谱,能够识别细胞亚群并定义其独特功能,因此已成为进一步解释疾病病理生理变化的强大工具。 [
8
] 然而,在组织解离过程中,单个细胞的空间分布及与其他细胞的相关性信息会丢失。空间转录组学(ST)的发明旨在结构性和空间层面上识别选定组织中的细胞类型及细胞间相互作用,从而弥补 scRNA‐seq 的不足。 [
9
] 将 scRNA‐Seq 与 ST 整合,相较于单独使用 scRNA‐Seq 或 ST,能够在损伤组织中实现病理的原位再现和可视化, [
10
] 并已应用于多种肿瘤疾病的研究。 [
11
] 因此,通过整合这两种技术,我们将能更好地理解 SAH 后 mLVs 的病理生理学。
In this study, we used scRNA‐seq and ST to comprehensively characterize the composition and alterations of different cell clusters in mLVs tissues after SAH in the mice model for the first time. After confirming that SAH could cause mLVs impairment, we then analyzed our sequencing data and discovered two significant factors, thrombospondin 1 (THBS1) and S100A6, were related to SAH prognosis. Finally, THBS1‐CD47 ligand‐receptor (L‐R) interaction was proved to induce meningeal lymphatic endothelial cells (mLECs) apoptosis through mediating STAT3/Bcl‐2 signaling, which might be the initial reason for mLVs injury after SAH.
在本研究中,我们首次利用单细胞 RNA 测序(scRNA-seq)和空间转录组学(ST)技术,全面描绘了小鼠模型在蛛网膜下腔出血(SAH)后脑膜淋巴管(mLVs)组织中不同细胞簇的组成与变化。在确认 SAH 可导致 mLVs 损伤的基础上,我们进一步分析测序数据,发现两个与 SAH 预后密切相关的关键因子——血小板反应蛋白 1(THBS1)和 S100A6。最终证实,THBS1-CD47 配体-受体(L-R)相互作用通过介导 STAT3/Bcl-2 信号通路诱导脑膜淋巴管内皮细胞(mLECs)凋亡,这可能是 SAH 后 mLVs 损伤的初始原因。
2. Results 2. 结果
2.1. Single‐Cell Transcriptomic Atlas of mLVs Alterations at Different Time Courses after SAH
2.1. SAH 后不同时间进程下 mLVs 变化的单细胞转录组图谱
To investigate whether mLVs are impaired after SAH, we used experimental SAH models by injecting autologous blood into the prechiasmatic cisternae. This model is ideal for exploring CNS lymphatic function because it does not demand complete exposure to neck anatomy, which can avoid direct impact on mLVs (Figure S1A, Supporting Information). Also, fluorescent beads were injected intra‐cisterna magna (i.c.m.) at various time courses after SAH modeling for further assessment of the drainage function of mLVs (Figure S1A, Supporting Information). The results presented a substantial decrease in beads drainage to the dCLNs as early as 3 h after SAH, and the drainage impairment remained until 72 h post‐injury (Figure
1B and Figure S1B, Supporting Information). As for the upstream path of dCLNs, decreased drainage and wounded structure also presented in mLVs at 24 and 72 h after SAH (Figure 1I and Figure S1C, Supporting Information).
为探究蛛网膜下腔出血(SAH)后脑膜淋巴管(mLVs)功能是否受损,我们采用向视交叉前池注入自体血液的方法建立实验性 SAH 模型。该模型因无需完全暴露颈部解剖结构,可避免对 mLVs 的直接损伤,是研究中枢神经系统淋巴功能的理想选择(图 S1A ,支持信息)。此外,在 SAH 建模后的不同时间点,通过小脑延髓池(i.c.m.)注射荧光微珠以进一步评估 mLVs 的引流功能(图 S1A ,支持信息)。结果显示,SAH 后最早 3 小时即可观察到微珠向深部颈淋巴结(dCLNs)的引流显著减少,且这种引流障碍持续至损伤后 72 小时(图
1B 和图 S1B ,支持信息)。在上游路径方面,SAH 后 24 小时和 72 小时,mLVs 也表现出引流减少及结构损伤(图 1I 和图 S1C ,支持信息)。
Figure 1. 图 1。
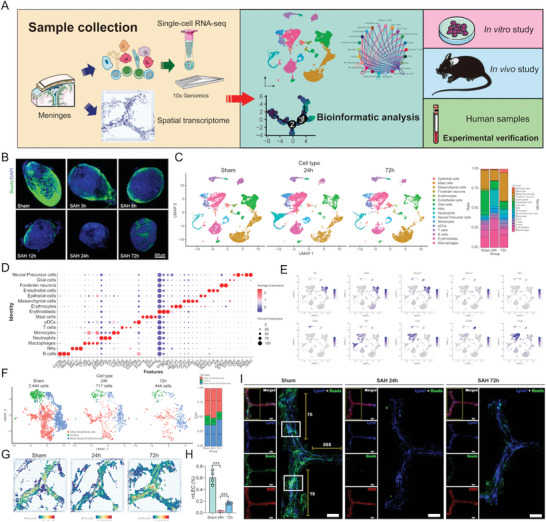
Single‐cell transcriptomic atlas of mLVs alterations at different time courses after SAH. A) Graphic overview of our study design. Mice dura matters were dissociated into single‐cell suspension and used for scRNA‐seq with 10× Genomics. Dura slides were processed to obtain spatial transcriptomics by 10× Genomics Visium. Integrated analysis of single‐cell transcriptome data and the following in vivo/vitro experiments were performed. B) Representative images of fluorescent beads accumulation in dCLNs at 3, 6, 12, 24, and 72 h post‐SAH. Scale bar: 200 µm. C) UMAP plots of 16 cell clusters with different colors (left) and bar charts of cluster proportion (right) in each group. D) Dot plots showing the average expression of known markers in 16 cell types. The dot size represents the percentage of cells expressing the indicated genes in each cell type. E) Expression levels of selected known marker genes across 25 536 unsorted cells illustrated in UMAP plots from both Sham and SAH dura in mice. F) High‐resolution UMAP plots of three re‐defined endothelial cell sub‐clusters in each group. G) Predicted spatial distribution of mLECs by RCTD. H) Flow cytometric analysis of mLECs percentage at different time courses after SAH, n = 4 per group, *** p < 0.001 versus Sham and 24 h by paired two‐tailed Student's t‐test. I) Representative confocal images of mLVs region at different time courses after SAH. An enlarged view of TS and SSS is shown on the right in each group. Solid boxes show the hotspots along the TS. Lyve1 (blue), Beads (green), and CD31 (red). Scale bar: 1000 µm.
SAH 后不同时间点 mLVs 变化的单细胞转录组图谱。A) 研究设计示意图。将小鼠硬脑膜组织解离为单细胞悬液,采用 10× Genomics 技术进行单细胞 RNA 测序。硬脑膜切片通过 10× Genomics Visium 平台处理获取空间转录组数据。整合分析单细胞转录组数据及后续体内/体外实验结果。B) SAH 后 3、6、12、24 及 72 小时荧光微球在 dCLNs 中聚集的代表性图像。比例尺:200 微米。C) 左图:16 个细胞簇的 UMAP 可视化(不同颜色标识);右图:各组细胞簇比例的条形图。D) 点图展示 16 种细胞类型中已知标志基因的平均表达量,点大小表示各细胞类型中表达特定基因的细胞百分比。E) UMAP 图中展示的小鼠假手术组与 SAH 组硬脑膜 25,536 个未分选细胞中选定已知标志基因的表达水平。F) 各组重新定义的内皮细胞亚群高分辨率 UMAP 图。G) RCTD 预测的 mLECs 空间分布。 H) SAH 后不同时间点 mLECs 百分比的流式细胞术分析,每组 n = 4,*** p < 0.001,与 Sham 组和 24 小时组相比,采用配对双尾 Student's t 检验。I) SAH 后不同时间点 mLVs 区域的代表性共聚焦图像。每组右侧显示 TS 和 SSS 的放大视图。实线框显示沿 TS 的热点区域。Lyve1(蓝色)、Beads(绿色)和 CD31(红色)。比例尺:1000 µm。
To comprehensively elucidate the cellular composition alteration of mLVs after SAH, we performed scRNA‐seq (10× Genomics) on mice in different groups (Sham group, SAH 24 h group, SAH 72 h group, ten mice per group) (Figure 1A). After excluding damaged or dead cells and putative cell doublets out of the original scRNA‐seq data and adjacent sequent analysis, a total of 25 536 cell transcriptomes of 30 mice were retained, in which 9891 cells were originated from the sham group, 8262 cells were from SAH 24 h group, and 7383 were cells from SAH 72 h group (Figure 1C). Then we normalized the gene expression referring to sequencing depth and mitochondrial read count and applied principal component analysis (PCA) based on highly variably expressed genes across cells. To correct the batch effect, we integrated scRNA‐seq data with the Seurat integration method.[
12
] Further, a unified uniform manifold approximation and projection (UMAP) embedding space and graph‐based clustering approach was used to identify cell clusters. Clusters were annotated with well‐known markers. The cells were classified into 16 major cell types (Figure 1C–E) including B cells marked by Cd79A, Cd79B, and Ms4a1,[
13
] NKs marked by Ncr1, Klrb1c, and Nkg7,[
14
] macrophages marked by C1qb, Cd68, Csf1r, and Mrc1,[
15
] neutrophils marked by S100α9, IL1β, and Csf3r,[
16
] monocytes marked by Ccr2 and Ly6c2,[
17
] T cells marked by Cd3d, Cd3e, and Cd247,[
18
] pDCs marked by Cd209a and Cd74,[
19
] mast cells marked by Fcer1a, Kit, and Ms4a2,[
20
] erythroblasts marked by Ttr, Enpp2, and Ptgds, erythrocyte marked by Car2, Hba‐a1, and Hbb‐bs,[
21
] mesenchymal cells marked by Col1a1, Col1a2, Col3a1, and DCN,[
13
] epithelial cells identified by the expression of Birc5 and Ube2c,[
22
] endothelial cell marked by Pecam1, Cdh5, and Kdr,[
13
] forebrain neuron marked by Gng13 and S100α5,[
23
] glial cell marked by S100b and Cdh2,[
24
] and neuralprecursor cells marked by Cdh2, Pde6g, and Gnb3.[
24
] Although all 16 major cell types were presented in mLVs both in sham and SAH groups (Figure 1C), the dynamic changes of each cell type were different. Meanwhile, given that mLECs originated from endothelial cells, we further performed clustering analysis for endothelial cells and obtained three sub‐clusters. This result showed that mLECs proportion became the least at 24 h after SAH (Figure 1F), indicating that at this time course, mLVs injury might be the severest.
为了全面阐明蛛网膜下腔出血(SAH)后 mLVs 的细胞组成变化,我们对不同组别(假手术组、SAH 24 小时组、SAH 72 小时组,每组十只小鼠)的小鼠进行了单细胞 RNA 测序(scRNA-seq,10× Genomics)(图 1A )。在排除原始 scRNA-seq 数据中的受损或死亡细胞及潜在的双细胞后,通过后续分析保留了 30 只小鼠共计 25,536 个细胞转录组,其中假手术组来源细胞 9,891 个,SAH 24 小时组 8,262 个,SAH 72 小时组 7,383 个(图 1C )。随后,我们根据测序深度和线粒体读数对基因表达进行标准化,并基于细胞间高变表达基因进行主成分分析(PCA)。为校正批次效应,采用 Seurat 整合方法对 scRNA-seq 数据进行整合 [
12
] 。进一步利用统一流形逼近与投影(UMAP)嵌入空间及基于图的聚类方法识别细胞簇,并通过已知标记物对细胞簇进行注释。 细胞被分类为 16 种主要类型(图 1C–E ),包括由 Cd79A、Cd79B 和 Ms4a1 标记的 B 细胞 [
13
] ,由 Ncr1、Klrb1c 和 Nkg7 标记的 NK 细胞 [
14
] ,由 C1qb、Cd68、Csf1r 和 Mrc1 标记的巨噬细胞 [
15
] ,由 S100α9、IL1β和 Csf3r 标记的中性粒细胞 [
16
] ,由 Ccr2 和 Ly6c2 标记的单核细胞 [
17
] ,由 Cd3d、Cd3e 和 Cd247 标记的 T 细胞 [
18
] ,由 Cd209a 和 Cd74 标记的 pDC 细胞 [
19
] ,由 Fcer1a、Kit 和 Ms4a2 标记的肥大细胞 [
20
] ,由 Ttr、Enpp2 和 Ptgds 标记的成红细胞,由 Car2、Hba‐a1 和 Hbb‐bs 标记的红细胞 [
21
] ,由 Col1a1、Col1a2、Col3a1 和 DCN 标记的间充质细胞 [
13
] ,通过 Birc5 和 Ube2c 表达鉴定的上皮细胞 [
22
] ,由 Pecam1、Cdh5 和 Kdr 标记的内皮细胞 [
13
] ,由 Gng13 和 S100α5 标记的前脑神经元 [
23
] ,由 S100b 和 Cdh2 标记的胶质细胞 [
24
] ,以及由 Cdh2、Pde6g 和 Gnb3 标记的神经前体细胞 [
24
] 。尽管在假手术组和 SAH 组的 mLVs 中均存在这 16 种主要细胞类型(图 1C ),但每种细胞类型的动态变化各不相同。 同时,考虑到 mLECs 来源于内皮细胞,我们进一步对内皮细胞进行了聚类分析,并获得了三个亚群。结果显示,SAH 后 24 小时时 mLECs 的比例降至最低(图 1F ),表明在这一时间点,mLVs 的损伤可能最为严重。
Similarly, flow cytometric analysis showed the population of CD45–CD31+LYVE1+ cells (mLECs) declined after SAH, and that the most serious injury happened 24 h post SAH (Figure 1H). Moreover, injured mice got lower modified Garcia scores, prolonged T
turn and T
total, prolonged deficits, and delayed recovery compared to the sham group in several neurological function tests, respectively (Figure S1D, Supporting Information). Taken together, these results demonstrated that SAH caused the injury of mLVs structure and drainage function, and elicited loss of neurological function in mice in the acute phase.
同样,流式细胞术分析显示,CD45 – CD31 + LYVE1 + 细胞(mLECs)群体在 SAH 后减少,且最严重的损伤发生在 SAH 后 24 小时(图 1H )。此外,与假手术组相比,损伤小鼠在多项神经功能测试中改良 Garcia 评分较低,T turn 和 T total, 延长,恢复延迟(图 S1D ,支持信息)。综上所述,这些结果表明 SAH 导致小鼠急性期 mLVs 结构和引流功能损伤,并引发神经功能丧失。
2.2. Deconvolution of Spatial Atlas and Visualization of Cell Type Interactions
2.2. 空间图谱反卷积与细胞类型互作可视化
As mentioned above, scRNA‐seq data lost the spatial characteristics of each cell. Therefore, we then combined ST to improve the integrity of sequencing data and reveal the spatial distribution of all cells. After hematoxylin and eosin (H&E) staining and brightfield imaging, mLVs slides were subjected to discover distinct histological features. Standard quality control and dimensionality reduction were performed using Seurat methods, and visualization was realized through t‐distributed Stochastic Neighbor Embedding (T‐SNE). Based on differentially expressed genes (DEGs) in each cluster, stepwise cell clustering was performed to construct an ST map. As was mentioned in a recent study, we similarly used RCTD software to deconvolute each tissue‐covering spot,[
25
] and further reconstructed the layered and segmented structure of sinus anatomy. The predicted localization of the 12 immune cell clusters confirmed their enrichment in specific regions (e.g., superior sagittal sinus [SSS], transverse sinus [TS], and hotspot). We mapped the ratios of each cell cluster within each spot as representative pie charts (scatter pies) both on the SSS and TS. Interestingly, distinct changes occurred in infiltrated immune cells into mLVs after injury. Compared to normal mLVs, infiltrated monocytes significantly increased and became the main group among all cell clusters 24 h after SAH. Although infiltrated macrophages were the most at 72 h with the majority subtype of M2, there were significantly more monocytes and M1 type macrophages gathering at the hotspot site (Figure
2A).
如上所述,scRNA-seq 数据丢失了每个细胞的空间特征。因此,我们随后结合 ST 技术以提高测序数据的完整性,并揭示所有细胞的空间分布。经过苏木精-伊红(H&E)染色和明场成像后,对 mLVs 切片进行观察以识别不同的组织学特征。使用 Seurat 方法进行标准质量控制和降维,并通过 t 分布随机邻域嵌入(t-SNE)实现可视化。基于每个簇中的差异表达基因(DEGs),逐步进行细胞聚类以构建 ST 图谱。如近期研究所述,我们同样采用 RCTD 软件对每个组织覆盖点进行反卷积, [
25
] 并进一步重建了窦解剖的分层和分段结构。12 个免疫细胞簇的预测定位证实了它们在特定区域(如上矢状窦[SSS]、横窦[TS]及热点区)的富集。我们将每个点内各细胞簇的比例以代表性饼图(散点饼图)形式映射在 SSS 和 TS 上。 有趣的是,损伤后浸润到 mLVs 中的免疫细胞发生了明显变化。与正常 mLVs 相比,SAH 后 24 小时,浸润的单核细胞显著增加,并成为所有细胞簇中的主要群体。尽管 72 小时时浸润的巨噬细胞最多,且以 M2 亚型为主,但在热点区域聚集的单核细胞和 M1 型巨噬细胞数量明显更多(图
2A )。
Figure 2. 图 2。
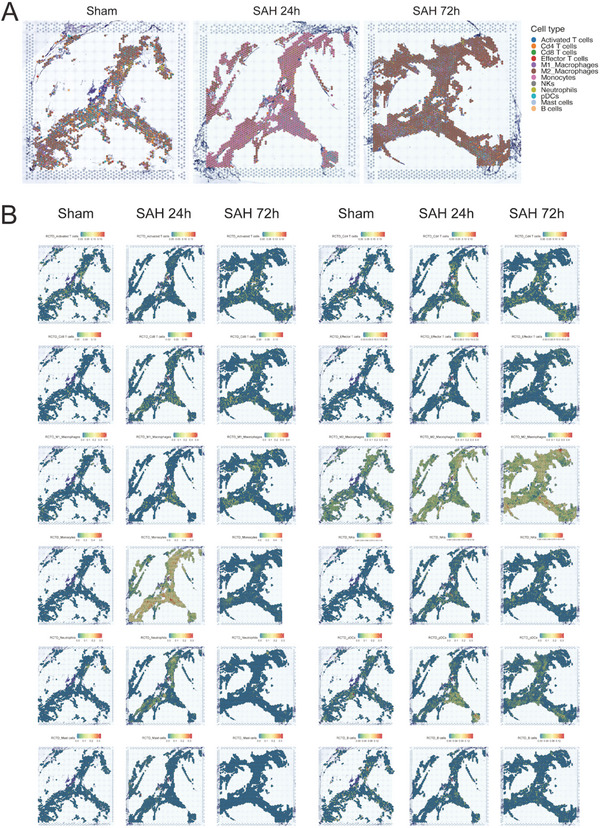
Deconvolution of spatial atlas of mLVs after SAH. A) Spatial scatter pie plots representing the proportion of different cell types from reference atlas within capture locations in mice dura at different time courses after SAH. B) Predicted spatial localization of immune cell types around mLVs by RCTD.
SAH 后 mLVs 空间图谱的去卷积分析。A) 空间散点饼图展示 SAH 后不同时间点小鼠硬脑膜捕获位点内参考图谱中不同细胞类型的比例。B) 通过 RCTD 预测的免疫细胞类型在 mLVs 周围的空间定位。
To further explore the alterations in the meningeal immune microenvironment, we attempted to map the scRNA‐seq data of each cell to ST to accurately reveal the infiltration of immune cells into mLVs after SAH. Immune cell infiltration was dominated by monocytes at 24 h and by macrophages at 72 h after SAH. Neutrophil infiltration peaked at 24 h and returned to normal level (sham) by 72 h. T cell infiltration gradually increased at different time courses after injury while B cell infiltration was relatively stable. NKs and pDC infiltration also continuously increased after injury (Figure 2B). These results reflected the cellular changes of mLVs in different stages as SAH progressed.
为深入探究脑膜免疫微环境的改变,我们尝试将单细胞 RNA 测序数据映射至空间转录组,以精确揭示蛛网膜下腔出血后免疫细胞向脑膜淋巴管的浸润情况。免疫细胞浸润以单核细胞在 24 小时占主导,而 72 小时后则以巨噬细胞为主。中性粒细胞浸润在 24 小时达到峰值,至 72 小时恢复至正常(假手术组)水平。T 细胞浸润在损伤后不同时间点逐渐增加,而 B 细胞浸润则相对稳定。自然杀伤细胞和浆细胞样树突状细胞的浸润在损伤后也持续增加(图 2B )。这些结果反映了蛛网膜下腔出血进展过程中脑膜淋巴管在不同阶段的细胞变化。
2.3. Cell‐to‐Cell Communication Analysis Revealed THBS1‐CD47 Ligand‐Receptor Pair Activation Associated with mLVs Impairment
2.3. 细胞间通讯分析揭示 THBS1-CD47 配体-受体对激活与脑膜淋巴管损伤相关
Given that immune inflammation had long been suggested as a key pathological mechanism involved in SAH pathogenesis,[
26
] and our results revealed the differences of infiltrated immune cells between the sham and SAH group, we wondered if the dynamic remodeling of immune microenvironment played an important role in mLVs impairment. Visualization of mLECs distribution prediction showed that mLECs damage was severest at 24 h after injury (Figure
3A). Therefore, we first investigated cell‐to‐cell communication among all cell types to visualize the overall situation. Through manifold learning and quantitative contrasts, CellChat identified differentially over‐expressed ligands and receptors between each cell cluster. In total, 4239 significant L‐R pairs were detected, which were further categorized into 2258 signaling pathways. Significant interactions were identified on the basis of a statistical test that randomly permuted the group labels of cells and then recalculated interaction probability.
鉴于免疫炎症长期以来被认为是参与 SAH 发病机制的关键病理机制, [
26
] 且我们的研究结果揭示了假手术组与 SAH 组之间浸润免疫细胞的差异,我们推测免疫微环境的动态重塑是否在 mLVs 损伤中起重要作用。mLECs 分布预测的可视化显示,损伤后 24 小时 mLECs 的损伤最为严重(图
3A )。因此,我们首先研究了所有细胞类型之间的细胞间通讯,以可视化整体情况。通过流形学习和定量对比,CellChat 识别了每个细胞簇之间差异过表达的配体和受体。总共检测到 4239 个显著的 L-R 对,这些被进一步分类为 2258 条信号通路。基于对细胞群标签进行随机置换后重新计算相互作用概率的统计检验,确定了显著的相互作用。
Figure 3. 图 3。
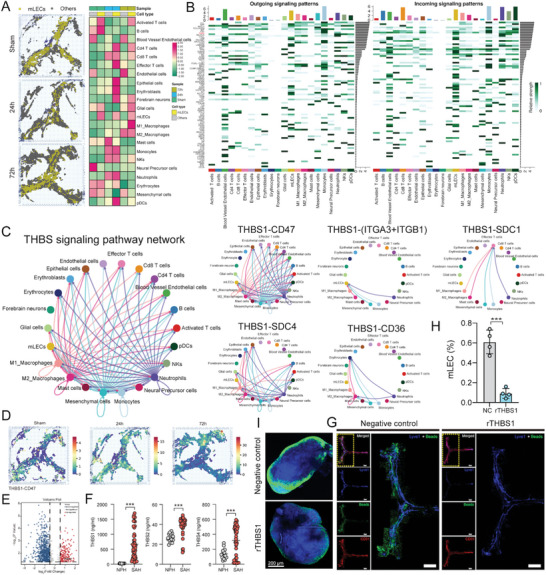
Intercellular communication networks in and around mLVs. A) Spatial visualization of immune cell infiltration based on mLECs distribution. B) Outgoing and incoming signal patterns among all meningeal cells. C) THBS signaling pathway network among all cell types. D) Spatial visualization of THBS1‐CD47 L‐R pair distribution by stLearn. E) Volcano plots showing that THBS1 is one of the upregulated DEGs in the SAH group compared to the NPH group identified by mass spectrometry. F) Quantification of THBS1, THBS2, and THBS4 expression in CSF samples from SAH patients compared to those from NPH patients by ELISA assay, *** p < 0.001 versus NPH by paired two‐tailed Student's t‐test. G) Representative confocal images of mLVs region of the negative control (NC) group and recombinant THBS1 protein treatment (rTHBS1) group. An enlarged view of TS and SSS is shown on the right in each group. Lyve1 (blue), Beads (green), and CD31 (red). Scale bar: 1000 µm. H) Flow cytometric analysis of mLECs percentage in NC group and rTHBS1 group, n = 4 per group, *** p < 0.001 versus NC by paired two‐tailed Student's t‐test. I) Representative images of beads accumulation in dCLNs of the NC group and rTHBS1 group. Scale bar: 200 µm.
脑膜淋巴管(mLVs)内及周围的细胞间通讯网络。A) 基于 mLECs 分布的免疫细胞浸润空间可视化。B) 所有脑膜细胞间的信号发出与接收模式。C) 各细胞类型间的 THBS 信号通路网络。D) 通过 stLearn 展示 THBS1-CD47 配体-受体对的空间分布。E) 火山图显示质谱鉴定中 THBS1 是 SAH 组相较于 NPH 组上调的差异表达基因之一。F) ELISA 检测 SAH 患者与 NPH 患者脑脊液样本中 THBS1、THBS2 和 THBS4 表达量的定量分析,*** p < 0.001(配对双尾 Student t 检验 vs NPH 组)。G) 阴性对照组(NC)与重组 THBS1 蛋白处理组(rTHBS1)mLVs 区域的代表性共聚焦图像。各组右侧显示 TS 和 SSS 的放大视图。Lyve1(蓝色)、微珠(绿色)和 CD31(红色)。比例尺:1000 µm。H) 流式细胞术分析 NC 组与 rTHBS1 组 mLECs 占比,每组 n=4,*** p < 0.001(配对双尾 Student t 检验 vs NC 组)。 I) NC 组和 rTHBS1 组 dCLNs 中珠子积聚的代表性图像。比例尺:200 µm。
Next, we set out to find the relationship between mLECs and other cell types. Several signals that contributed most to the outgoing or incoming signaling patterns of mLECs were identified (Figure 3B). According to the findings above, mLVs injury was most severe at 24 h after injury and monocytes were the dominant infiltrated cells at this time course, indicating that monocytes and mLECs interaction might play crucial roles in mLVs dysfunction. Therefore, by focusing on the communication between mLECs and monocytes, we found that THBS signaling pathways were especially actively involved in all signaling patterns. We further explored the potential L‐R pairs based on THBS signaling working on mLECs and calculated the contribution of these L‐R pairs. Among them, the L‐R pair “THBS1‐CD47” exhibited the strongest interaction in cell‐to‐cell communication (Figure 3C and Figure S2A,B, Supporting Information). To further assess the spatial distribution of the THBS1‐CD47 L‐R pair, we used stLearn,[
27
] a technology especially used to analyze spatial communication, which could combine the spatial information of cells with all the predicted communication to discover the regional specificity of L‐R pairs. L‐R co‐expression enrichment among neighbor spots and cell‐type density enrichment were integrated to find hotspots within a tissue, where interactions between cell types were most likely to occur. The result showed that the THBS1‐CD47 L‐R pair significantly activated at 24 h after SAH (Figure 3D).
接下来,我们着手探究 mLECs 与其他细胞类型之间的关系。识别出对 mLECs 传出或传入信号模式贡献最大的若干信号(图 3B )。基于上述发现,损伤后 24 小时 mLVs 损伤最为严重,此时单核细胞是主要的浸润细胞群,表明单核细胞与 mLECs 的相互作用可能在 mLVs 功能障碍中起关键作用。因此,通过聚焦 mLECs 与单核细胞间的通讯,我们发现 THBS 信号通路在所有信号模式中尤为活跃。我们进一步挖掘了作用于 mLECs 的潜在 L-R 配对,并计算了这些配对的贡献度。其中,"THBS1-CD47" L-R 对在细胞间通讯中展现出最强的相互作用(图 3C 和图 S2A,B ,支持信息)。 为进一步评估 THBS1-CD47 L-R 对的空间分布,我们采用了 stLearn 技术( [
27
] ),这是一种专门用于分析空间通讯的技术,能够将细胞的空间信息与所有预测的通讯相结合,以发现 L-R 对的区域特异性。通过整合邻近点之间的 L-R 共表达富集和细胞类型密度富集,我们识别出组织内最可能发生细胞类型间互作的热点区域。结果显示,THBS1-CD47 L-R 对在 SAH 后 24 小时显著激活(图 3D )。
2.4. THBS1 Impaired Meningeal Lymphatic Drainage and Exacerbated Neurological Dysfunction
2.4. THBS1 损害脑膜淋巴引流并加剧神经功能障碍
After discovering that the THBS1‐CD47 L‐R pair was potentially associated with mLVs injury, we then would like to assess the impact of THBS1 after SAH. We checked endogenous THBS1 alteration after SAH by using our previous mass spectrometry proteomics data from ProteomeXchange Consortium (PXD030593). Differentially expressed proteins in human CSF of SAH patients measured by mass spectrometry indicated significant up‐regulation of THBS1 protein (we used CSF samples from normal pressure hydrocephalus (NPH) patients as the control group, Figure 3E). Given that the human THBS family had three subtypes, including THBS1, THBS2, and THBS4,[
28
] we used an ELISA kit to again test the expression of all THBS subtypes in CSF samples of SAH patients to verify if THBS1 was the key THBS member. As we expected, although THBS2 and THBS4 levels also elevated, expression of THBS1 increased most after SAH compared to the NPH group (Figure 3F and Table S1, Supporting Information). Also, higher THBS1 level was strongly connected with poorer clinical outcome in SAH patients (Figure 6F).
在发现 THBS1-CD47 配体-受体对可能与脑膜淋巴管(mLVs)损伤相关后,我们进一步评估了 SAH 后 THBS1 的影响。通过分析先前提交至 ProteomeXchange 联盟(PXD030593)的质谱蛋白质组学数据,我们检测了 SAH 后内源性 THBS1 的变化。质谱测定的人 SAH 患者脑脊液差异表达蛋白显示 THBS1 蛋白显著上调(以正常压力脑积水(NPH)患者脑脊液作为对照组,图 3E )。鉴于人类 THBS 家族包含 THBS1、THBS2 和 THBS4 三种亚型 [
28
] ,我们采用 ELISA 试剂盒复测 SAH 患者脑脊液中所有 THBS 亚型的表达以验证 THBS1 是否为关键成员。如预期所示,虽然 THBS2 和 THBS4 水平亦有升高,但相较于 NPH 组,SAH 后 THBS1 的表达增幅最为显著(图 3F 及附表 S1 ,支持信息)。此外,较高的 THBS1 水平与 SAH 患者较差的临床预后密切相关(图 6F )。
Figure 6. 图 6。
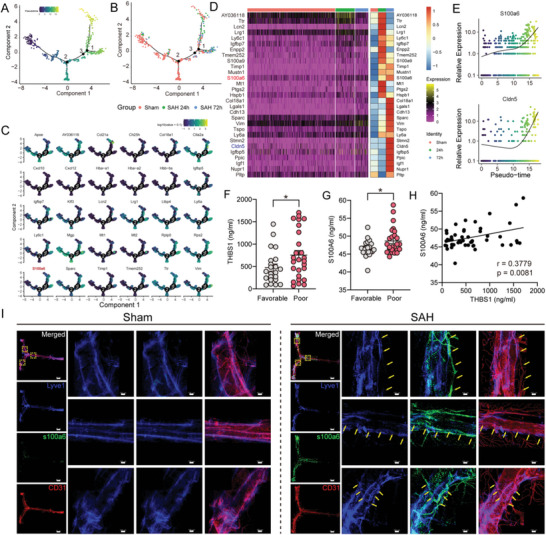
Cell trajectory analysis shows the evolution of mLECs. A) Labeling mLECs with real grouping information based on pseudotime trajectory map. B) Pseudotime trajectory map. C) Trajectory of representative genes in mLECs. D) Heatmap showing the top 30 pseudotime‐related genes. E) Representative two pseudo‐time‐related genes (S100α6 and Cldn5) based on q values. F,G) Increased expression of THBS1 and S100A6 are associated with poor prognosis, n = 48, *p < 0.05 by paired two‐tailed Student's t‐test. H) Linear relationship between THBS1 and S100A6 expression, r = 0.3779, p = 0.0081 by Spearman's rank test. H) Representative confocal images of mLVs region of sham and SAH 24 h group. Enlarged view of selected region in the merged photo (yellow dotted box) are listed on the right in each group. Lyve1 (blue), S100α6 (green), and CD31 (red). Yellow arrows indicate where mLVs lie. Scale bar: 800 µm for the holistic view and 100 µm for the enlarged view.
细胞轨迹分析展示了 mLECs 的演化过程。A) 基于伪时间轨迹图,用实际分组信息标记 mLECs。B) 伪时间轨迹图。C) mLECs 中代表性基因的轨迹。D) 显示前 30 个与伪时间相关基因的热图。E) 基于 q 值的两个代表性伪时间相关基因(S100α6 和 Cldn5)。F,G) THBS1 和 S100A6 表达增加与不良预后相关,n=48,*p<0.05(配对双尾 Student's t 检验)。H) THBS1 与 S100A6 表达之间的线性关系,r=0.3779,p=0.0081(Spearman 秩检验)。H) 假手术组和 SAH 24 小时组 mLVs 区域的代表性共聚焦图像。每组右侧列出了合并照片中选定区域(黄色虚线框)的放大视图。Lyve1(蓝色)、S100α6(绿色)和 CD31(红色)。黄色箭头指示 mLVs 所在位置。比例尺:整体视图 800 µm,放大视图 100 µm。
We then performed in vivo experiments to explore the role of THBS1 in mLVs injury after SAH. First, low‐dosage mouse recombinant THBS1 protein was delivered into mice cisterna magna to observe mLVs function changes. Compared to negative control (NC) mice, recombinant THBS1 (rTHBS1) protein treatment caused less beads aggregation in mLVs and dCLNs (Figure 3G,I and Figure S3A, Supporting Information) and decreased mLECs number of mLVs (Figure 3H). Second, we constructed THBS1‐overexpressing mice through the delivery of AAV‐THBS1 into mice cisterna magna, along with cultivating THBS1‐KO mice for further research. Compared to AAV‐control, mLVs injury was even worse with AAV‐THBS1 treatment after SAH (Figure
4A–C and Figure S3B, Supporting Information). Conversely, THBS1‐KO partly reversed mLVs damage caused by SAH (Figure 4H–J and Figure S3C, Supporting Information). Also, neurological function tests presented similar results. Overexpression of THBS1 aggravated neurological deficits caused by SAH, while THBS1 knockout improved neurofunction (Figure 4D–G,K–N). What is more, consistent with the results of THBS1 knockout, mLVs and neurofunction impairment was reversed as well after blockage of THBS1 with THBS1‐antibody (Figure
5A–G and Figure S3D, Supporting Information). Collectively, all these results proved that up‐regulated THBS1 caused mLVs structure and drainage function damage after SAH, thereby exacerbating neurobehavioral dysfunction.
随后,我们进行了体内实验以探究 THBS1 在 SAH 后 mLVs 损伤中的作用。首先,将低剂量小鼠重组 THBS1 蛋白注入小鼠小脑延髓池,观察 mLVs 功能变化。与阴性对照(NC)小鼠相比,重组 THBS1(rTHBS1)蛋白处理导致 mLVs 和 dCLNs 中珠子聚集减少(图 3G,I 和图 S3A ,支持信息),并降低了 mLVs 中 mLECs 的数量(图 3H )。其次,我们通过向小鼠小脑延髓池递送 AAV-THBS1 构建了 THBS1 过表达小鼠,同时培养 THBS1-KO 小鼠用于进一步研究。与 AAV 对照相比,SAH 后 AAV-THBS1 处理使 mLVs 损伤更为严重(图
4A–C 和图 S3B ,支持信息)。相反,THBS1-KO 部分逆转了 SAH 引起的 mLVs 损伤(图 4H–J 和图 S3C ,支持信息)。此外,神经功能测试也呈现相似结果。THBS1 的过表达加剧了 SAH 导致的神经功能缺损,而 THBS1 敲除则改善了神经功能(图 4D–G,K–N )。 此外,与 THBS1 敲除结果一致,使用 THBS1 抗体阻断 THBS1 后,脑膜淋巴管(mLVs)和神经功能损伤同样得到逆转(支持信息中的图
5A–G 和图 S3D )。综上所述,这些结果均证实,SAH 后上调的 THBS1 导致了脑膜淋巴管结构和引流功能损伤,从而加剧了神经行为功能障碍。
Figure 4. 图 4。
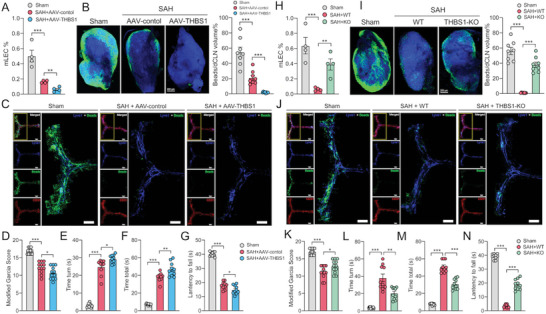
Validation of THBS1 impacting mLVs function after SAH. SAH modeling and behavioral tests were performed 4 weeks after the AAV injection. A) Flow cytometric analysis of mLECs percentage in Sham, AAV‐control, and AAV‐THBS1 group at 72 h post‐SAH, n = 4 per group, **p < 0.01, ***p < 0.001 by paired two‐tailed Student's t‐test. B) Representative images and quantification of beads accumulation in dCLNs of Sham, AAV‐control, and AAV‐THBS1 group at 72 h post‐SAH, each data point represents an average of the 2 dCLNs from one individual mouse, n = 8 per group, **p < 0.01, ***p < 0.001 by paired two‐tailed Student's t‐test. Scale bar: 200 µm. C) Representative confocal images of mLVs region of Sham, AAV‐control, and AAV‐THBS1 group at 72 h post‐SAH. An enlarged view of TS and SSS is shown on the right in each group. Lyve1 (blue), Beads (green), and CD31 (red). Scale bar: 1000 µm. D) Modified Garcia test, E) time turn, F) time total, and G) wire hanging test at 72 h after SAH revealed AAV‐THBS1 delivery aggravated short‐term neurological function compared with Sham or AAV‐control group, n = 10–12 per group. *p < 0.05; **p < 0.01, ***p < 0.001 by paired two‐tailed Student's t‐test. H) Flow cytometric analysis of mLECs percentage in Sham, SAH‐WT, and THBS1‐KO group at 24 h post‐SAH, n = 4 per group, **p < 0.01, ***p < 0.001 by paired two‐tailed Student's t‐test. I) Representative images and quantification of beads accumulation in dCLNs in sham, SAH‐WT, and THBS‐KO group at 24 h post‐SAH, each data point represents an average of the 2 dCLNs from one individual mouse. n = 8 per group, **p < 0.01 by paired two‐tailed Student's t‐test. Scale bar: 200 µm. J) Representative confocal images of mLVs region of Sham, SAH‐WT, and THBS1‐KO group at 24 h post‐SAH. An enlarged view of TS and SSS is shown on the right in each group. Lyve1 (blue), Beads (green), and CD31 (red). Scale bar: 1000 µm. K) Modified Garcia test, L) time turn, M) time total, and N) wire hanging test at 24 h after SAH revealed THBS1 knockout improved short‐term neurological function compared with Sham or WT‐SAH group. n = 10–12 per group. *p < 0.05; **p < 0.01, ***p < 0.001 by paired two‐tailed Student's t‐test.
验证 THBS1 对 SAH 后 mLVs 功能的影响。AAV 注射后 4 周进行 SAH 建模及行为学测试。A) SAH 后 72 小时,假手术组、AAV 对照组与 AAV-THBS1 组中 mLECs 比例的流式细胞术分析,每组 n=4,**p < 0.01,***p < 0.001(配对双尾 Student's t 检验)。B) SAH 后 72 小时各组 dCLNs 中微球蓄积的代表性图像与定量,每个数据点代表单只小鼠双侧 dCLNs 的平均值,每组 n=8,**p < 0.01,***p < 0.001(配对双尾 Student's t 检验)。比例尺:200 µm。C) SAH 后 72 小时各组 mLVs 区域的共聚焦显微镜代表性图像,右侧为各组 TS 和 SSS 区域的放大视图(Lyve1 标记为蓝色,微球为绿色,CD31 为红色)。比例尺:1000 µm。D) 改良 Garcia 评分、E) 转向时间、F) 总时间及 G) 悬丝试验显示,SAH 后 72 小时 AAV-THBS1 组较假手术组或 AAV 对照组短期神经功能恶化,每组 n=10–12。*p < 0.05;**p < 0.01,***p < 0.001(配对双尾 Student's t 检验)。 H) 假手术组、SAH-WT 组及 THBS1-KO 组在 SAH 后 24 小时 mLECs 百分比的流式细胞术分析,每组 n=4,**p < 0.01,***p < 0.001(配对双尾 Student's t 检验)。I) SAH 后 24 小时假手术组、SAH-WT 组及 THBS-KO 组 dCLNs 中微珠积累的代表性图像及定量,每个数据点代表一只小鼠 2 个 dCLNs 的平均值。每组 n=8,**p < 0.01(配对双尾 Student's t 检验)。比例尺:200 µm。J) SAH 后 24 小时假手术组、SAH-WT 组及 THBS1-KO 组 mLVs 区域的代表性共聚焦图像。各组右侧显示 TS 和 SSS 的放大视图。Lyve1(蓝色)、微珠(绿色)及 CD31(红色)。比例尺:1000 µm。K) 改良 Garcia 测试,L) 转身时间,M) 总时间,及 N) 悬挂测试在 SAH 后 24 小时显示,与假手术组或 WT-SAH 组相比,THBS1 敲除改善了短期神经功能。每组 n=10–12。*p < 0.05;**p < 0.01,***p < 0.001(配对双尾 Student's t 检验)。
Figure 5. 图 5。
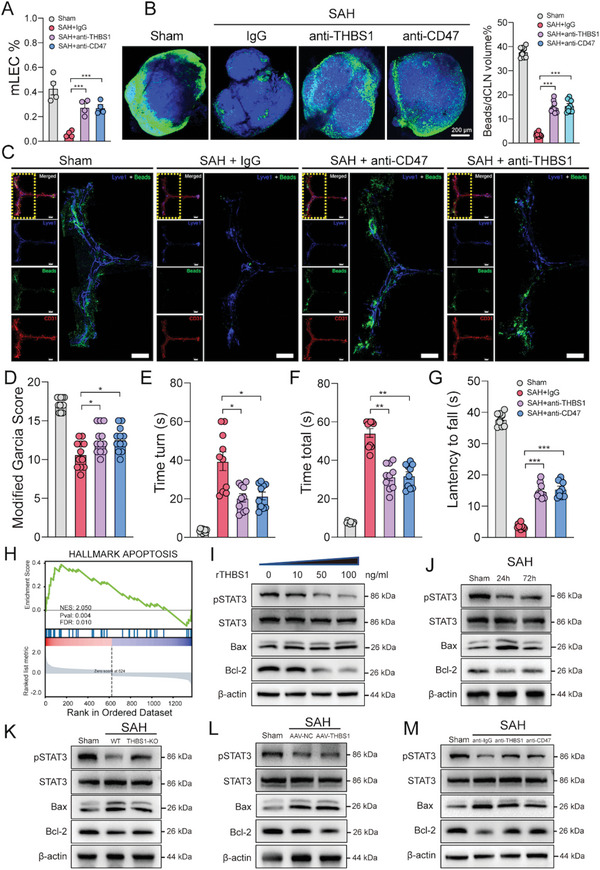
Disturbing THBS1‐CD47 interaction promoted mLVs restoration via inhibiting STAT3/BCL2‐mediated apoptosis in mLECs. A) Flow cytometric analysis of mLECs percentage in sham, SAH + igG, SAH + anti‐CD47, and SAH + anti‐THBS1 group at 24 h post‐SAH, n = 4 per group, ***p < 0.001 by paired two‐tailed Student's t‐test. B) Representative images and quantification of beads accumulation in dCLNs of Sham, SAH + igG, SAH + anti‐CD47, and SAH + anti‐THBS1 group at 24 h post‐SAH, each data point represents an average of the 2 dCLNs from one individual mouse, n = 8 per group, ***p < 0.001 by paired two‐tailed Student's t‐test. Scale bar: 200 µm. C) Representative confocal images of mLVs region of Sham, SAH + igG, SAH + anti‐CD47, and SAH + anti‐THBS1 group at 24 h post‐SAH. An enlarged view of TS and SSS is shown on the right in each group. Lyve1 (blue), Beads (green), and CD31 (red). Scale bar: 1000 µm. D) Modified Garcia test, E) time turn, F) time total, and G) wire hanging test at 24 h after SAH revealed anti‐CD47 and anti‐THBS1 therapy improved short‐term neurological function compared with Sham or SAH + igG group. n = 10–12 per group. *p < 0.05; **p < 0.01, ***p < 0.001 by paired two‐tailed Student's t‐test. H) GSEA showed apoptosis pathway was activated in mLECs after SAH. I) Representative immunoblot images showing effects of rTHBS1 (100 ng mL−1) treatment on pSTAT3 and Bcl‐2 inhibition in primary mLECs. J–M) Representative immunoblot images of STAT3, pSTAT3, Bax, and Bcl‐2 in different groups.
干扰 THBS1-CD47 相互作用通过抑制 STAT3/BCL2 介导的 mLECs 凋亡促进 mLVs 恢复。A) SAH 后 24 小时,假手术组、SAH+igG 组、SAH+抗 CD47 组及 SAH+抗 THBS1 组中 mLECs 比例的流式细胞术分析,每组 n=4,***p < 0.001(配对双尾 Student t 检验)。B) SAH 后 24 小时,假手术组、SAH+igG 组、SAH+抗 CD47 组及 SAH+抗 THBS1 组 dCLNs 中微球蓄积的代表性图像与定量,每个数据点代表单只小鼠双侧 dCLNs 的平均值,每组 n=8,***p < 0.001(配对双尾 Student t 检验)。比例尺:200 µm。C) SAH 后 24 小时各组 mLVs 区域的共聚焦显微镜代表性图像,右侧为各组 TS 和 SSS 区域的放大视图。Lyve1(蓝色)、微球(绿色)及 CD31(红色)。比例尺:1000 µm。D) 改良 Garcia 评分,E) 转向时间,F) 总时间,G) 悬丝试验显示,与假手术组或 SAH+igG 组相比,抗 CD47 和抗 THBS1 治疗在 SAH 后 24 小时改善了短期神经功能。每组 n=10–12。 *p < 0.05;**p < 0.01,***p < 0.001,采用配对双尾 Student t 检验。H) GSEA 分析显示 SAH 后 mLECs 中凋亡通路被激活。I) 代表性免疫印迹图像展示 rTHBS1(100 ng/mL)处理对原代 mLECs 中 pSTAT3 及 Bcl-2 抑制的影响。J–M) 不同组别中 STAT3、pSTAT3、Bax 和 Bcl-2 的代表性免疫印迹图像。
2.5. Disturbing THBS1‐CD47 Interaction Promoted mLVs Restoration via Inhibiting STAT3/BCL2‐Mediated Apoptosis in mLECs
2.5. 干扰 THBS1-CD47 相互作用通过抑制 STAT3/BCL2 介导的 mLECs 凋亡促进 mLVs 修复
Previous studies reported that THBS1 could inhibit endothelial cell proliferation and chemotaxis by engaging its receptors CD47.[
29
] Similarly, our results predicted that the THBS1‐CD47 L‐R pair played an important role in mLVs impairment and proved the injurious effect of THBS1 after SAH. We therefore moved on to discover CD47 function. Blockage of CD47 with antibody attenuated meningeal lymphatic dysfunction compared to igG treatment at 24 h post‐SAH (Figure 5A–C and Figure S3D, Supporting Information). Accordantly, the neurological function also improved significantly (Figure 5D–G).
先前研究报道,THBS1 可通过与其受体 CD47 结合抑制内皮细胞增殖和趋化作用 [
29
] 。类似地,我们的结果预测 THBS1-CD47 配体-受体对在 SAH 后脑膜淋巴管损伤中起重要作用,并证实了 THBS1 的损害效应。因此,我们进一步探究了 CD47 的功能。与 IgG 处理相比,SAH 后 24 小时用抗体阻断 CD47 可减轻脑膜淋巴功能障碍(图 5A–C 和图 S3D ,支持信息)。相应地,神经功能也显著改善(图 5D–G )。
Next, to further investigate the underlying mechanism of THBS1‐CD47 interaction‐induced injury, GSEA analysis was performed.[
30
] The result indicated that the apoptosis pathway in mLECs was significantly activated 24 h after injury (Figure 5H). Tunel staining indicated that apoptosis was significantly activated after SAH. Overexpression of THBS1 further promoted the apoptosis of mLVs while blockage of THBS1 showed decreased Lyve1+Tunel+ cells (Figure S5A, Supporting Information). Co‐cultivation of human monocytes and LECs in vitro confirmed that the content of THBS1 in the supernatant of the co‐culture system was significantly increased under the stimulation of hemoglobin (150 µm), indicating that monocytes are one of the sources of THBS1 secretion after SAH (Figure S5B, Supporting Information). Flow cytometric analysis revealed that the apoptosis of LECs in the co‐culture system pretreated with THBS1 neutralizing antibody is significantly reduced compared to the control group (Figure S5C,D, Supporting Information). However, the precise mechanism of mLVs injury remains unknown.
接下来,为进一步探究 THBS1-CD47 相互作用诱导损伤的潜在机制,我们进行了 GSEA 分析。 [
30
] 结果显示,损伤后 24 小时 mLECs 中的凋亡通路被显著激活(图 5H )。Tunel 染色表明 SAH 后凋亡显著激活。THBS1 的过表达进一步促进了 mLVs 的凋亡,而阻断 THBS1 则显示 Lyve1 + Tunel + 细胞减少(图 S5A ,支持信息)。体外共培养人单核细胞与 LECs 证实,在血红蛋白(150 µm)刺激下,共培养系统上清液中 THBS1 含量显著增加,表明单核细胞是 SAH 后 THBS1 分泌的来源之一(图 S5B ,支持信息)。流式细胞术分析显示,与对照组相比,经 THBS1 中和抗体预处理的共培养系统中 LECs 的凋亡显著减少(图 S5C,D ,支持信息)。然而,mLVs 损伤的确切机制仍不清楚。
Studies had demonstrated that THBS1‐CD47 interaction induced endothelial cell apoptosis through disturbing VEGF/VEGFR2 signaling, which was essential for endothelial development.[
29
] Also, VEGFR2 inhibition was found to promote apoptosis via suppressing STAT3/Bcl‐2 signaling in osteosarcoma.[
31
] Therefore, we hypothesized if THBS1‐CD47 interaction could medicate mLECs apoptosis via inhibiting STAT3/Bcl‐2 signaling. First, co‐immunoprecipitation (COIP) confirmed the interaction between THBS1 and CD47 in LECs (Figure S4A, Supporting Information). Second, along with increased expression of Bax, expression of pSTAT3 and Bcl‐2 were significantly decreased after rTHBS1treatment in primary mLECs in a dose‐dependent manner (Figure 5I), indicating that rTHBS1 treatment promoted mLECs apoptosis by repressing the activation of the STAT3/Bcl‐2 signaling pathway. Finally, in SAH mice mLVs samples, the level of pSTAT3 and Bcl‐2 peaked at 24 h and then decreased at 72 h after SAH, which was consistent with GSEA results (Figure 5J). THBS1 knockout or blockage of THBS1/CD47 could all significantly increase pSTAT3 and Bcl‐2 expression (Figure 5K,M), whereas THBS1‐overexpression presented the opposite effect (Figure 5L). Taken together, all these findings verified that the THBS1‐CD47 L‐R pair promoted mLECs apoptosis via STAT3/Bcl‐2 signaling, and inhibition of this interaction might help alleviate mLVs injury.
研究表明,THBS1-CD47 相互作用通过干扰 VEGF/VEGFR2 信号传导诱导内皮细胞凋亡,而该信号对内皮发育至关重要。 [
29
] 此外,发现抑制 VEGFR2 可通过抑制 STAT3/Bcl-2 信号通路促进骨肉瘤细胞凋亡。 [
31
] 因此,我们推测 THBS1-CD47 相互作用是否可能通过抑制 STAT3/Bcl-2 信号通路介导 mLECs 凋亡。首先,共免疫沉淀(COIP)证实了 LECs 中 THBS1 与 CD47 的相互作用(图 S4A ,支持信息)。其次,在原代 mLECs 中,随着 Bax 表达的增加,rTHBS1 处理后 pSTAT3 和 Bcl-2 的表达呈剂量依赖性显著下降(图 5I ),表明 rTHBS1 处理通过抑制 STAT3/Bcl-2 信号通路的激活促进了 mLECs 凋亡。最后,在 SAH 小鼠 mLVs 样本中,pSTAT3 和 Bcl-2 的水平在 SAH 后 24 小时达到峰值,随后在 72 小时下降,这与 GSEA 结果一致(图 5J )。 THBS1 基因敲除或阻断 THBS1/CD47 相互作用均能显著增加 pSTAT3 和 Bcl-2 的表达(图 5K,M ),而过表达 THBS1 则呈现相反效果(图 5L )。综上所述,这些发现证实 THBS1-CD47 配体-受体对通过 STAT3/Bcl-2 信号通路促进 mLECs 凋亡,抑制该相互作用可能有助于减轻 mLVs 损伤。
2.6. Pseudotime Analysis Revealed mLECs Alteration Trace after SAH
2.6. 伪时序分析揭示 SAH 后 mLECs 的变化轨迹
Additionally, we performed pseudotime analysis of all mLECs in different samples to reveal mLECs change trajectory during the EBI stage after SAH (Figure
6A). After labeling mLECs with real grouping information, we found that the damage pattern of mLECs 24 h after SAH was consistent with the findings above. However, unexpectedly, mLECs at 72 h seemed to possess completely different cell states or characteristics compared to the sham group and 24 h group (Figure 6B). Considering that mLVs impairment was partly improved at 72 h after SAH (Figure 1I), we wondered if self‐repairing mechanisms of mLECs or other cell‐to‐cell interactions functioned at this time course, which was worth the further efforts and exploration. DEG analysis revealed the most over‐expressed genes after SAH (Figure 6C,D). Along the trajectory, the expression of S100α6 and Cldn5 gradually increased (Figure 6E). Previous research reported that S100A6 impaired lymphatic endothelial cells tight junction and increased the transendothelial migration of neutrophils in breast cancer,[
32
] indicating that S100α6 might be a potential mLECs damage biomarker after SAH.
此外,我们对不同样本中的所有 mLECs 进行了伪时间分析,以揭示 SAH 后 EBI 阶段 mLECs 的变化轨迹(图
6A )。在用真实分组信息标记 mLECs 后,我们发现 SAH 后 24 小时 mLECs 的损伤模式与上述发现一致。然而,出乎意料的是,与假手术组和 24 小时组相比,72 小时的 mLECs 似乎具有完全不同的细胞状态或特征(图 6B )。考虑到 SAH 后 72 小时 mLVs 损伤部分改善(图 1I ),我们推测此时可能有 mLECs 的自我修复机制或其他细胞间相互作用在发挥作用,这值得进一步研究和探索。差异表达基因(DEG)分析揭示了 SAH 后表达最上调的基因(图 6C,D )。沿轨迹方向,S100α6 和 Cldn5 的表达逐渐增加(图 6E )。先前研究报道,S100A6 会损害淋巴管内皮细胞的紧密连接并增加中性粒细胞在乳腺癌中的跨内皮迁移, [
32
] 这表明 S100α6 可能是 SAH 后 mLECs 损伤的潜在生物标志物。
We then performed immunofluorescent staining of mLVs to identify S100α6 changes after SAH. As expected, there was a marked increase in meningeal S100α6 expression and more Lyve1+S100α6+ positive cells 24 h after injury while Lyve1+ cells decreased compared to the uninjured group (Figure 6I). Conversely, the expression of S100α6 was also significantly reduced when THBS1 was knocked out (Figure S6, Supporting Information). Also, through the ELISA assay, we confirmed increased expression of S100A6 in CSF samples of SAH patients compared to the control. Prognostic analysis suggested higher expression of S100A6 was related to a worse prognosis (Figure 6G and Table S1, Supporting Information). Meanwhile, the expression of S100A6 and THBS1 had a linear relationship (Figure 6H), which indicated that both of them might be appropriate predictors after SAH. In summary, our data illustrated that S100α6 might be a novel biomarker of meningeal lymphatic injury, which further supplemented the gaps in the field of mLVs annotation.
随后,我们对脑膜淋巴管(mLVs)进行了免疫荧光染色,以观察 SAH 后 S100α6 的变化。如预期所示,损伤 24 小时后,脑膜中 S100α6 表达显著增加,且 Lyve1 + 与 S100α6 + 双阳性细胞增多,而未损伤组的 Lyve1 + 细胞则有所减少(图 6I )。相反,当 THBS1 基因敲除时,S100α6 的表达也显著降低(图 S6 ,支持信息)。此外,通过 ELISA 检测,我们证实 SAH 患者脑脊液样本中 S100A6 的表达较对照组升高。预后分析显示,S100A6 的高表达与不良预后相关(图 6G 及表 S1 ,支持信息)。同时,S100A6 与 THBS1 的表达呈线性关系(图 6H ),表明二者可能均为 SAH 后合适的预测指标。总之,我们的数据表明 S100α6 可能是脑膜淋巴损伤的新型生物标志物,进一步填补了 mLVs 注释领域的空白。
3. Discussion 3. 讨论
In this study, we integrated scRNA‐seq and ST to systematically delineate the cellular, molecular, and spatial alterations of injured mLVs after SAH for the first time. Analysis of DEGs between sham and SAH groups showed that gene transcription in each cell cluster and immune microenvironment underwent widespread changes after SAH. Cell‐to‐cell communication revealed secreted protein THBS1 binding to receptor CD47 visibly activated in mLECs after SAH. We hypothesized that activation of THBS1‐CD47 interaction after SAH promoted mLECs apoptosis by inhibiting the STAT3/Bcl‐2 signaling pathway, thereby impacting mLVs function. We verified our hypothesis through both in vivo/vitro experiments. Our research demonstrated that multi‐omics profiling can provide more comprehensive information from different perspectives, enabling better resolution of cellular composition, decipherment of immune cell infiltration, and explanation of the underlying pathological mechanism of mLVs after SAH.
本研究首次整合单细胞 RNA 测序(scRNA-seq)与空间转录组(ST)技术,系统描绘了蛛网膜下腔出血(SAH)后损伤脑膜淋巴管(mLVs)在细胞、分子及空间层面的变化。差异表达基因(DEGs)分析显示,假手术组与 SAH 组相比,各细胞簇及免疫微环境中的基因转录均发生广泛改变。细胞间通讯分析揭示,SAH 后 mLECs 中分泌蛋白 THBS1 与受体 CD47 的结合显著激活。我们提出假设:SAH 后 THBS1-CD47 相互作用的激活通过抑制 STAT3/Bcl-2 信号通路促进 mLECs 凋亡,进而影响 mLVs 功能。该假设通过体内外实验得到验证。研究表明,多组学分析能从不同维度提供更全面的信息,有助于更精准解析细胞组成、破译免疫细胞浸润模式并阐明 SAH 后 mLVs 的潜在病理机制。
SAH is usually caused by a rupture of intracranial aneurysms. A mass of blood suddenly flushes into SAS through the ruptured aneurysm and results in subsequent brain injury.[
33
] On one hand, massive aggregation of blood in SAS lead to rapid elevation of ICP.[
34
] On the other hand, the breakdown of red blood cells releases several intracellular components, such as hemoglobin, into CSF, which can directly exert neurotoxic effects.[
33
] These two processes are devastating and are the predominant causes of EBI after SAH.[
35
] Although several endogenous mechanisms have been discovered, like erythrophagocytosis, to have the potential of clearing hemoglobin or other neurotoxicants, these mechanisms are quite easy to get overloaded or dampened.[
36
] Therefore, it is urgent and meaningful to further discover the pathological mechanism after SAH and find out a much more effective approach for neurotoxicant removal.
SAH 通常由颅内动脉瘤破裂引起。大量血液通过破裂的动脉瘤突然涌入 SAS,导致后续的脑损伤。 [
33
] 一方面,SAS 中血液的大量聚集导致颅内压迅速升高。 [
34
] 另一方面,红细胞的破裂释放出多种细胞内成分,如血红蛋白,进入脑脊液,可直接产生神经毒性作用。 [
33
] 这两个过程极具破坏性,是 SAH 后 EBI 的主要原因。 [
35
] 尽管已发现几种内源性机制,如红细胞吞噬作用,具有清除血红蛋白或其他神经毒性物质的潜力,但这些机制极易过载或受抑制。 [
36
] 因此,进一步探索 SAH 后的病理机制并寻找更有效的神经毒性物质清除方法具有紧迫性和重要意义。
The CNS is traditionally considered an immune‐privileged organ, mainly due to the lack of direct communication or connection with the peripheral immune system.[
37
] However, the existence of lymphatic vessels (LVs) in the dura, known as mLVs, has been confirmed in both mice and humans.[
5
,
38
] Given that mLVs drain CSF and interstitial fluid (ISF) into dCLNs, it is reasonable to surmise that these vessels are involved in clearing waste, regulating the flow of CSF, and maintaining brain homeostasis under physiological conditions.[
5
,
39
] The discovery of mLVs and subsequent studies have improved our understanding of several CNS diseases including Alzheimer's disease, traumatic brain injury, and subdural hematomas.[
6
,
40
] As aforementioned, Chen et al. found that extravasated erythrocytes from SAS were drained into dCLNs via mLVs.[
3b
] In addition, ablation of mLVs exacerbated EBI symptoms in mice after SAH.[
41
] These discoveries identified mLVs as an important path for intracranial waste clearing and a promising therapeutic target for accelerating erythrocyte drainage after SAH. Nevertheless, the cellular architecture and the underlying gene regulatory features of mLVs post‐SAH have not been fully characterized. Our current project intended to apply scRNA‐Seq combined with ST to reveal the underlying mechanism of mLVs injury after SAH.
传统上,中枢神经系统(CNS)被视为免疫豁免器官,主要因其缺乏与外周免疫系统的直接交流或连接。 [
37
] 然而,硬脑膜中存在被称为 mLVs 的淋巴管(LVs),这一现象已在小鼠和人类中得到证实。 [
5
,
38
] 鉴于 mLVs 将脑脊液(CSF)和组织间液(ISF)引流至深部颈淋巴结(dCLNs),有理由推测这些血管在生理条件下参与清除废物、调节脑脊液流动及维持大脑稳态。 [
5
,
39
] mLVs 的发现及后续研究增进了我们对包括阿尔茨海默病、创伤性脑损伤和硬膜下血肿在内的多种中枢神经系统疾病的理解。 [
6
,
40
] 如前所述,Chen 等人发现蛛网膜下腔(SAS)溢出的红细胞通过 mLVs 被引流至 dCLNs。 [
3b
] 此外,mLVs 的消融加剧了小鼠在蛛网膜下腔出血(SAH)后的早期脑损伤(EBI)症状。 [
41
] 这些发现确立了 mLVs 作为颅内废物清除的重要途径,并成为加速 SAH 后红细胞引流的一个有前景的治疗靶点。 然而,SAH 后 mLVs 的细胞结构及其潜在的基因调控特征尚未完全阐明。我们当前的项目旨在应用 scRNA-Seq 结合 ST 技术,揭示 SAH 后 mLVs 损伤的潜在机制。
MLVs mainly lie in the dura, which is the outermost of the three layers of meninges that cover the brain and spinal cord. Unlike the adjacent brain parenchyma, a wide repertoire of immune cells is located in the dura and takes part in the constitution of meningeal immunity, which is concerned with the immune surveillance of CNS.[
42
] Meningeal immunity has recently come under the spotlight owing to the discovery of mLVs drainage function. Immune cell population alteration, immune‐inflammatory signaling pathway activation, and reactive substance generation are activated in the brain parenchyma after SAH. Crosstalk between immune dysfunction and hyperactivation of inflammatory signals aggravates the devastating consequences of brain injury and cerebral vasospasm. Presently, research on immune inflammation after SAH mainly focuses on injury in brain parenchyma. However, the changes in the immune microenvironment in dura remain unclear. In this study, we discovered that around mLVs, neutrophils, monocytes, and macrophages were markedly infiltrated at 24 h, and pDCs and T cells gradually increased and peaked at 72 h post‐SAH. These results were consistent with the trend of immune cell infiltration in the brain parenchyma after SAH, proposing a possibility that meningeal immune cells could directly migrate to parenchyma through mLVs after SAH.
MLVs 主要位于硬脑膜中,硬脑膜是覆盖大脑和脊髓的三层脑膜的最外层。与邻近的脑实质不同,硬脑膜中存在多种免疫细胞,参与构成脑膜免疫系统,该系统与中枢神经系统的免疫监视有关。 [
42
] 由于发现 MLVs 的引流功能,脑膜免疫最近成为研究热点。SAH 后,脑实质中免疫细胞群变化、免疫炎症信号通路激活及反应性物质生成被触发。免疫功能障碍与炎症信号过度激活之间的相互作用加剧了脑损伤和脑血管痉挛的破坏性后果。目前,关于 SAH 后免疫炎症的研究主要集中在脑实质损伤上。然而,硬脑膜内免疫微环境的变化尚不清楚。本研究中,我们发现 SAH 后 24 小时,MLVs 周围中性粒细胞、单核细胞和巨噬细胞显著浸润,而 pDCs 和 T 细胞逐渐增多并在 72 小时达到峰值。 这些结果与 SAH 后脑实质中免疫细胞浸润的趋势一致,提出了脑膜免疫细胞在 SAH 后可能通过 mLVs 直接迁移至脑实质的可能性。
THBS1 is a member of the thrombospondin protein family.[
43
] It is involved in tissue remodeling and with implications for tumor formation, wound healing, and embryonic development.[
44
] THBS1 exerts different effects by binding to different cellular receptors.[
45
] Previous studies have shown that THBS1 is highly expressed in lymphoma,[
46
] breast cancer,[
47
] and oral squamous carcinoma,[
48
] promoting tumor cell adhesion, proliferation, apoptosis, invasion, and metastasis. We discovered that THBS1 expression was elevated in CSF after SAH, and its level was associated with the prognosis of SAH patients, indicating that THBS1 might be a potential biomarker of SAH. Also, THBS1 is the first identified endogenous inhibitor of angiogenesis.[
29a
] It plays only a minor role in developmental angiogenesis, but it is an important regulator of pathological angiogenesis in the adult. Through interacting with its receptors CD36 and CD47, THBS1 can block endothelial cell proliferation and chemotaxis stimulated by fibroblast growth factor‐2 and VEGF and further induce endothelial cell apoptosis.[
29a
] By combining scRNA‐Seq and ST analysis, our present research predicted that mLECs apoptosis induced by THBS1‐CD47 interaction might be the key mechanism of mLVs dysfunction after SAH. After several in vivo/vitro experiments, we proved this hypothesis and further discovered that the downstream STAT3/Bcl‐2 signaling pathway participated in mLECs apoptosis.
THBS1 是血小板反应蛋白家族的一员。 [
43
] 它参与组织重塑,并与肿瘤形成、伤口愈合及胚胎发育密切相关。 [
44
] THBS1 通过与不同细胞受体结合发挥多样化作用。 [
45
] 既往研究表明,THBS1 在淋巴瘤、 [
46
] 乳腺癌、 [
47
] 及口腔鳞状细胞癌中高表达, [
48
] 促进肿瘤细胞黏附、增殖、凋亡、侵袭和转移。我们发现 SAH 后脑脊液中 THBS1 表达升高,其水平与 SAH 患者预后相关,提示 THBS1 可能是 SAH 的潜在生物标志物。此外,THBS1 是首个被鉴定的内源性血管生成抑制剂。 [
29a
] 在发育性血管生成中作用微弱,但对成体病理性血管生成具有重要调控作用。通过与受体 CD36 和 CD47 相互作用,THBS1 能阻断成纤维细胞生长因子-2 和 VEGF 刺激的内皮细胞增殖与趋化,并进一步诱导内皮细胞凋亡。 [
29a
] 通过整合单细胞 RNA 测序与空间转录组分析,本研究预测 SAH 后 THBS1-CD47 相互作用诱导的 mLECs 凋亡可能是 mLVs 功能障碍的关键机制。经系列体内外实验验证后,我们证实了这一假说,并进一步发现下游 STAT3/Bcl-2 信号通路参与了 mLECs 的凋亡过程。
Up to now, there is no research on mLVs injury after SAH. Our research attempted to use a multi‐omics study to describe the atlas of mLVs after SAH, to find potential injury mechanisms, and to mine more biological markers. S100α6 protein belongs to A group of the S100 protein family of Ca2+ binding proteins.[
49
] As an intracellular protein, S100α6 has been implicated in the regulation of several cellular functions, such as proliferation, apoptosis, cytoskeleton dynamics, and cellular response, to different stress factors.[
50
] Previous studies revealed S100α6 impaired lymphatic endothelial cells tight junction and increased trans‐endothelial migration of neutrophils in breast cancer.[
32
] Meanwhile, the dosage of serum S100α6 might aid in diagnosis in oncology.[
50b
] Our study identified increased expression of S100α6 both in mLVs and human CSF after SAH and its correlation with poor prognosis, indicating S100α6 might be a valuable biomarker.
迄今为止,尚无关于蛛网膜下腔出血(SAH)后脑膜淋巴管(mLVs)损伤的研究。我们的研究尝试通过多组学方法描绘 SAH 后 mLVs 的图谱,探索潜在的损伤机制,并挖掘更多生物标志物。S100α6 蛋白属于 S100 钙结合蛋白家族的 A 组,作为一种细胞内蛋白,它参与调控多种细胞功能,包括增殖、凋亡、细胞骨架动态变化及对不同应激因子的细胞反应。先前研究表明,S100α6 会损害淋巴管内皮细胞的紧密连接,并增加乳腺癌中中性粒细胞的跨内皮迁移。同时,血清 S100α6 的剂量可能有助于肿瘤学的诊断。我们的研究发现,SAH 后 mLVs 和人脑脊液中 S100α6 表达增加,且与不良预后相关,表明 S100α6 可能是一个有价值的生物标志物。
There are several limitations to our study. First, our study provided strong data to support an association among SAH, mLVs, and THBS1/CD47 L‐R pair because THBS1 or CD47 blockage improves mLVs in SAH mice. However, if these beneficial effects are directly mediated by mLECs THBS1/CD47 signal is not known. Since THBS1/CD47 L‐R pair affects many cell types, it is also possible that changes in mLVs and mLECs are indirectly influenced by THBS1/CD47 in other cell types. Second, the possible sources of THBS1 or S100A6 after SAH should be discussed.
我们的研究存在若干局限性。首先,本研究提供了强有力的数据支持蛛网膜下腔出血(SAH)、脑膜淋巴管(mLVs)与 THBS1/CD47 配体-受体对之间的关联,因为阻断 THBS1 或 CD47 能改善 SAH 小鼠的 mLVs 功能。然而,这些有益效应是否直接由 mLECs 中的 THBS1/CD47 信号介导尚不明确。鉴于 THBS1/CD47 配体-受体对影响多种细胞类型,mLVs 和 mLECs 的变化也可能间接受到其他细胞类型中 THBS1/CD47 的影响。其次,需探讨 SAH 后 THBS1 或 S100A6 的可能来源。
In conclusion, for the first time, we illustrated a landscape of spatial and cellular alterations of mLVs at an early stage after SAH. We discovered significant mLECs population change with the remodeling of the surrounding immune microenvironment. Meanwhile, overexpression of THBS1 and interaction between THBS1‐CD47 led to mLECs apoptosis possibly through STAT3/Bcl‐2 signaling. Taken together, our findings may provide new perspectives for the following SAH‐related research and treatment strategies for SAH intervention based on the improvement of impaired mLVs.
总之,我们首次描绘了 SAH 早期阶段脑膜淋巴管(mLVs)在空间和细胞层面的改变全景。发现伴随周围免疫微环境重塑,mLECs 群体发生显著变化。同时,THBS1 的过表达及其与 CD47 的相互作用可能通过 STAT3/Bcl-2 信号通路导致 mLECs 凋亡。综上所述,我们的发现可能为后续 SAH 相关研究及基于改善受损 mLVs 的 SAH 干预治疗策略提供新视角。
4. Experimental Section 4. 实验部分
Clinical Sample Collection and Patient Information
临床样本采集与患者信息
Patients with aneurysmal SAH were diagnosed by computed tomography (CT) and digital subtraction angiograph (DSA) performed within 24 h after admissions. The inclusion and exclusion criteria were consistent with the previous study.[
51
] All CSF samples of SAH patients were collected before coiling or clipping through a lumbar puncture or lumbar drainage (LD) or external ventricular drainage (EVD) within 48 h after SAH. All CSF sample collection and patient information were consistent with the previous report.[
3a
]
动脉瘤性蛛网膜下腔出血(SAH)患者通过入院 24 小时内进行的计算机断层扫描(CT)和数字减影血管造影(DSA)确诊。纳入与排除标准与先前研究一致。 [
51
] 所有 SAH 患者的脑脊液(CSF)样本均在出血后 48 小时内通过腰椎穿刺、腰椎引流(LD)或脑室外引流(EVD)在弹簧圈栓塞或夹闭手术前采集。所有 CSF 样本采集及患者信息均与先前报告一致。 [
3a
]
Mass Spectrometry of Human CSF
人类脑脊液的质谱分析
The previous mass spectrometry proteomics data from ProteomeXchange Consortium (PXD030593) was used for downstream analysis. Used the hiplot tool to present relevant gene expression in the form of a volcano plot.[
52
]
先前来自 ProteomeXchange 联盟(PXD030593)的质谱蛋白质组学数据被用于下游分析。使用 hiplot 工具以火山图形式展示相关基因表达。 [
52
]
ELISA Assay ELISA 检测
The concentrations of human CSF THBS1 (EK0899, Boster Biotech, Wuhan), THBS2 (EK0642, Boster Biotech, Wuhan), THBS4 (CSB‐EL023490HU, Cusabio Biotech, Wuhan), and S100A6 (CSB‐EL020634RA, Cusabio Biotech, Wuhan) were determined using ELISA kits. Human CSF samples (100 µL) were used for the analysis. All procedures were performed according to the manufacturer's instructions.
采用 ELISA 试剂盒测定人脑脊液中 THBS1(EK0899,Boster Biotech,武汉)、THBS2(EK0642,Boster Biotech,武汉)、THBS4(CSB‐EL023490HU,Cusabio Biotech,武汉)和 S100A6(CSB‐EL020634RA,Cusabio Biotech,武汉)的浓度。分析使用 100 微升人脑脊液样本,所有操作均按照制造商说明书进行。
Animals 动物
Specific pathogen‐free, C57BL/6 male mice (6–8 weeks old) were purchased from Shanghai SLAC Laboratory Animal Co. Ltd. For AAV injection, C57BL/6 mice were injected with AAV‐Ctrl, AAV‐THBS1 at 6 weeks of age, when the weight was ≈20 g. SAH modeling and behavioral tests were performed 4 weeks after the AAV injection. C57BL/6 mice were injected into with recombinant mice THBS1 protein or vehicle (PBS) via cisterna magna 1 day in advance before the sacrifice. THBS1−/‐ mice were established on a C57BL/6 background and purchased from Gempharmatech Co., Ltd (#KO‐T014586, NCBI:21 825, MGI:98 737). Mice were housed in the animal facility with controlled habituation and temperature, on 12‐h light versus dark cycles, and fed with regular rodents' chow and sterilized tap water ad libitum. Mice were allowed to accommodate for 2 weeks before experimental procedures.
无特定病原体(SPF 级)的 C57BL/6 雄性小鼠(6-8 周龄)购自上海斯莱克实验动物有限责任公司。对于腺相关病毒(AAV)注射,C57BL/6 小鼠在 6 周龄、体重约 20 克时分别注射 AAV-Ctrl 或 AAV-THBS1。AAV 注射 4 周后进行蛛网膜下腔出血(SAH)模型构建及行为学测试。在处死前 1 天,C57BL/6 小鼠通过小脑延髓池注射重组小鼠 THBS1 蛋白或对照溶剂(PBS)。THBS1 −/‐ 小鼠基于 C57BL/6 背景构建,购自集萃药康生物科技有限公司(品系编号 KO-T014586,NCBI:21825,MGI:98737)。小鼠饲养于环境温湿度可控的动物设施中,采用 12 小时明暗循环,自由摄食常规啮齿类饲料及灭菌自来水。所有小鼠在实验操作前均适应环境 2 周。
Induction of SAH SAH 诱导
The prechiasmatic SAH mice model was created as previously described.[
53
] In brief, after induction of anesthesia with 1% pentobarbital, the head was fixed in a stereotactic apparatus (RWD Life Science Co., Ltd). With a midline incision, the skin overlying the anterior skull was opened. A burr hole was drilled into the skull 4.5 mm anterior to the bregma with a caudal angel of 40° using a 0.9‐mm drill. Blood (60 µL) was withdrawn from a C57BL/6 WT blood donor and injected over a 10‐s period with a 26‐gauge needle advanced through the burr hole at a 40° angle until reaching the base of the skull. For antibody injection, a mixture of 10 µg mL−1 CD47 antibody (Invitrogen, 16‐0479‐85) or THBS1 antibody (Genetex, GTX21823) or IgG (Invitrogen, 14‐4714‐85) with 60 µL autologous blood was injected into the prechiasmatic cisternae. The needle was left in place for 5 min before retraction to avoid backflow. Mice that underwent the same procedure, without blood and antibody/IgG injections, served as the sham group.
视交叉前蛛网膜下腔出血(SAH)小鼠模型的建立方法如前所述 [
53
] 。简而言之,使用 1%戊巴比妥麻醉后,将小鼠头部固定于立体定位仪(RWD Life Science Co., Ltd)。沿中线切开前颅骨表面皮肤,用 0.9 毫米钻头在距前囟 4.5 毫米处以 40 度尾倾角钻孔。从 C57BL/6 野生型供血鼠抽取 60 微升血液,通过 26 号针头以 40 度角经骨孔推进至颅底,在 10 秒内完成注射。抗体注射时,将 10 微克/毫升的 CD47 抗体(Invitrogen, 16-0479-85)、THBS1 抗体(Genetex, GTX21823)或 IgG(Invitrogen, 14-4714-85)与 60 微升自体血液混合后注入视交叉前池。针头留置 5 分钟后缓慢退出以防止反流。假手术组小鼠接受相同操作但不注射血液及抗体/IgG。
Intra‐Cisterna Magna Injections
小脑延髓池内注射
Mice were anesthetized by intraperitoneal (i.p.) injection with 1% pentobarbital. The skin of the neck was shaved and cleaned with iodine and 70% ethanol, and an ophthalmic solution (erythromycin ointment) was placed on the eyes to prevent drying. Stereotaxic frames were used to secure the mouse's head, and a midline incision was made in the skin. The muscle layers were retracted and the cisterna magna exposed. Using a Hamilton syringe (coupled to a 33‐gauge needle), the volume of the desired solution was injected into the CSF‐filled cisterna magna compartment. For THBS1 or Control AAV delivery experiments, 2 µL of artificial CSF containing 1013 genome copies per mL of AAV9‐THBS1, or AAV‐Con (AAV9, adeno‐associated virus serotype 9; purchased from PackGene Biotech, Guangzhou), were injected into the cisterna magna CSF at a rate of 2 µL min−1. For recombinant protein experiments, recombinant mice THBS1 protein (150 µg kg−1, R&D Systems, 7859‐TH‐050, USA) or vehicle (PBS) were injected into cisterna magna CSF at a rate of 2 µL min−1. For the beads experiments, 2 µL of FluoSpheres carboxylate 0.5 µm‐beads 505/515 (Invitrogen, F8813) were injected at a rate of 2 µL min−1. The needle was inserted into the cisterna magna through the retracted muscle in order to prevent backflow upon needle removal. The neck skin was then sutured and allowed to recover on a heating pad until fully awake.
小鼠通过腹腔注射 1%戊巴比妥钠麻醉。颈部皮肤剃毛并用碘伏和 70%乙醇消毒,眼部涂抹眼科用药(红霉素软膏)以防干燥。使用立体定位仪固定小鼠头部,沿中线切开皮肤。分离肌肉层暴露小脑延髓池。采用汉密尔顿注射器(连接 33 号针头)将指定体积的溶液注入充满脑脊液的小脑延髓池腔。在 THBS1 或对照 AAV 递送实验中,以 2 微升/分钟的速率向小脑延髓池脑脊液内注射 2 微升含 10 13 基因组拷贝/毫升的 AAV9-THBS1 或 AAV-Con(AAV9,腺相关病毒血清型 9,购自广州派真生物)的人工脑脊液。重组蛋白实验中,以 2 微升/分钟的速率注射重组小鼠 THBS1 蛋白(150 微克/千克 −1 ,R&D Systems 货号 7859-TH-050,美国)或载体(PBS)。微球实验中,以 2 微升/分钟的速率注射 2 微升 0.5 微米羧基荧光微球 505/515(Invitrogen 货号 F8813)。 将针头通过回缩的肌肉插入小脑延髓池,以防止拔针时回流。随后缝合颈部皮肤,并在加热垫上恢复直至完全清醒。
Neurological Scores and Behavioral Testing
神经学评分和行为测试
Neurological scores and all behavioral assessments of each group were carried out at similar and relatively fixed time during daylight hours in a blinded fashion.
各组神经学评分及所有行为评估均在白天相似且相对固定的时间内以盲法进行。
Neurological Scores 神经学评分
The Modified Garcia (ranging from 0 to 18) was conducted to evaluate short‐term neurological function. The scoring system was comprised of six sub‐tests including response capacity, alertness, coordination, motor skills, complex movements, and coordination.[
54
] A blinded investigator performed the neurological examination, which was scored 24 and 72 h after SAH. A higher score represented a better neurological function.
采用改良 Garcia 评分(范围 0~18 分)评估短期神经功能。该评分系统包括反应能力、警觉性、协调性、运动技能、复杂动作和协调性六个子测试。 [
54
] 由盲法研究者进行神经学检查,分别在 SAH 后 24 小时和 72 小时评分。评分越高,神经功能越好。
Pole Test 杆测试
A pole test was performed 24 and 72 h postoperatively. Animals are placed on top of a 50‐ to 55‐cm vertical pole with a diameter of 8 to 10 mm and trained to turn around and descend the pole. The scoring started when the animal initiated the turning movement. The time to make a complete 180° turn (i.e., T
turn) and latency to reach the ground (i.e., T
total) were recorded. If the animal cannot turn but instead descends with a lateral body position, then T
total was usually attributed to T
turn. When an animal makes a turn, descends halfway, and falls, the recorded time stops when the animal reaches the floor. The maximum time to perform the test was set to 60 s. Mice were trained three times daily for 3 days before the start of testing.
术后 24 小时和 72 小时进行杆测试。将动物置于直径 8 至 10 毫米、高 50 至 55 厘米的垂直杆顶端,训练其转身并沿杆下行。评分自动物开始转身动作时启动。记录完成 180 度转身所需时间(即 T turn )及到达地面的延迟时间(即 T total )。若动物无法转身而以侧身姿势下行,则通常将 T total 归为 T turn 。当动物完成转身、下行中途坠落时,记录时间止于其触地瞬间。测试最长时限设为 60 秒。正式测试前,小鼠每日训练三次,持续三天。
Wire Hanging Test 钢丝悬挂试验
Wire Hanging is a simple test that evaluates grip strength, balance, and endurance. Mice were trained to suspend their body by holding on to a single wire stretched between two posts 50 to 60 cm above the ground. To prevent the animal from using all four paws, the hind limbs were gently covered with adhesive tape. Between the two posts was a pillow to avoid injury when the mice fell. “Latency to fall” was the primary end point used to assess motor performance. The maximum time to perform the test was set to 60 s. Mice were trained three times daily for 3 days before the start of testing.
钢丝悬挂是一项评估抓握力、平衡性和耐力的简单测试。训练小鼠通过抓住一根离地 50 至 60 厘米、横跨两柱之间的单根钢丝来悬吊身体。为防止动物使用所有四爪,后肢被轻柔地用胶带覆盖。两柱之间放置了一个垫子,以防小鼠跌落时受伤。“跌落潜伏期”是评估运动表现的主要终点指标。测试的最长时间设定为 60 秒。在测试开始前,小鼠每天接受三次训练,持续三天。
Tissue Collection 组织采集
Mice were euthanized with isoflurane and dCLNs were dissected under the microscope. Subsequently drop‐fixed in 4% paraformaldehyde (PFA) for 2 h at 4 °C and then the CUBIC clearance protocol was performed as described below in the dCLN clearance methods sections.[
55
] For meningeal whole‐mount collection, mice were transcardially perfused with 20 mL cold 1× PBS, then skin and muscle were stripped from the outer skull, and the skullcap was removed with surgical scissors and fixed in 2% PFA for 12 h at 4 °C. Then the meninges (dura mater and arachnoid mater) were carefully dissected from the skullcaps with Dumont #5 forceps (Fine Science Tools). Meningeal whole‐mounts were then moved to PBS and 0.05% azide at 4 °C until further use.
小鼠通过异氟烷安乐死后,在显微镜下解剖出 dCLNs。随后用 4%多聚甲醛(PFA)在 4°C 下滴固定 2 小时,然后按照下文 dCLN 透明化方法部分所述的 CUBIC 透明化流程进行处理。 [
55
] 对于脑膜全片采集,小鼠经心脏灌注 20 mL 冰冷的 1×PBS,然后剥离头皮和肌肉,用手术剪取下颅盖,置于 2% PFA 中 4°C 固定 12 小时。随后使用 Dumont #5 镊子(精细科学工具)从颅盖上小心分离脑膜(硬脑膜和蛛网膜)。脑膜全片随后转移至含 0.05%叠氮化钠的 PBS 中,4°C 保存待用。
dCLN Clearance dCLN 透明化
Consistent with the previous report,[
41
] dCLN clearance was performed following the published CUBIC protocol with modifications.[
55
] In brief, nodes were incubated in 50% reagent 1 (prepared 1:1 with dH2O) for 1 day at 37 °C, shaking, with DAPI (1:1000). Nodes were then transferred to reagent 1 for 1 day at 37 °C, shaking, with DAPI (1:1000). Nodes were washed two times in PBS + 0.01% sodium azide for 2 h and overnight with DAPI (1:1000) at 37 °C. Then nodes were incubated with 50% reagent 2 (prepared 1:1 with dH2O) for 1 day at 37 °C with DAPI (1:1000). Finally, nodes were incubated with reagent 2 for 1 day at 37 °C. Nodes were placed in eight well chambers (155 411, Thermo Fisher) with mineral oil and imaged with confocal microscopy.
与先前报告一致, [
41
] dCLN 清除按照已发表的 CUBIC 方案及修改步骤进行。 [
55
] 简言之,淋巴结在 50%试剂 1(以 dH2O 按 1:1 配制)中于 37°C 震荡孵育 1 天,加入 DAPI(1:1000)。随后将淋巴结转移至试剂 1 中,于 37°C 震荡孵育 1 天,加入 DAPI(1:1000)。淋巴结在 PBS + 0.01%叠氮化钠中洗涤两次,每次 2 小时,并于 37°C 与 DAPI(1:1000)过夜孵育。接着,淋巴结在 50%试剂 2(以 dH2O 按 1:1 配制)中于 37°C 与 DAPI(1:1000)孵育 1 天。最后,淋巴结在试剂 2 中于 37°C 孵育 1 天。将淋巴结置于含矿物油的八孔室(155 411,赛默飞世尔)中,使用共聚焦显微镜成像。
Immunofluorescence, Imaging, and Quantification
免疫荧光、成像与定量分析
For immunofluorescence staining, meningeal whole‐mounts in PBS and 0.05% azide were blocked with either 2% donkey serum or 2% goat serum, 1% bovine serum albumin, 0.1% triton, 0.05% tween‐20, and 0.05% sodium azide in PBS for 1.5 h at room temperature. This blocking step was followed by incubation with appropriate dilutions of primary antibodies: Rat anti‐Lyve1(Santa Cruze, sc‐65647, 1:200), Goat anti‐CD31 (R&D Systems, AF3628, 1:100), and Rabbit anti‐S100α6 (Abcam, ab181975, 1:200) in the same solution used for blocking overnight at 4 °C. Meningeal whole‐mounts or brain tissue sections were then washed three times for 10 min at room temperature in PBS and 0.05% tween‐20, followed by incubation with the appropriate donkey Alexa Fluor 405 or 488 or 594 anti‐rat, ‐goat,‐rabbit (Thermo Fisher Scientific, 1:1000) for 2 h at RT in the same solution used for blocking. For Tunel staining, apoptotic cells were detected according to the manufacturer's instructions using the apoptotic kit (Beyotime Biotech, C1090, China). The whole mounts were then washed three times for 10 min at RT in PBS. The tissue was then transferred to PBS and mounted with ProLong Gold antifade reagent (Invitrogen, P36934) on glass slides with coverslips. Slide preparations were stored at 4 °C and imaged using a Lecia DMI8 confocal microscope and LAS AF software (Leica Microsystems) within 1 week of staining. Quantitative analysis of the acquired images was performed using Fiji software. For lymph nodes, the percent volume of microbead coverage in cleared dCLN was assessed by creating a 3D reconstruction of the node and then calculating the volume covered by beads divided by the total volume of the node using Fiji. The right and the left dCLN percent volume were averaged together for each mouse. For assessment of mLV coverage and complexity, images of the meningeal whole‐mounts were acquired using a confocal microscope and Fiji was used for quantifications. The entire meningeal whole‐mount was traced and used for quantification. The percent area coverage of Lyve1 was used to determine the coverage of the lymphatic vessels. When applicable, the same images were used to assess the percentage of field coverage by Lyve1−CD31+ vessels. All meningeal whole mounts used for the quantification of lymphatic morphology were imaged with identical confocal settings and Fiji parameters.
进行免疫荧光染色时,将 PBS 和 0.05%叠氮化物中的脑膜全片用 2%驴血清或 2%山羊血清、1%牛血清白蛋白、0.1% Triton、0.05% Tween-20 及 0.05%叠氮化钠的 PBS 溶液在室温下封闭 1.5 小时。封闭后,用适当稀释的一抗于 4°C 过夜孵育:大鼠抗 Lyve1(Santa Cruze, sc-65647, 1:200)、山羊抗 CD31(R&D Systems, AF3628, 1:100)及兔抗 S100α6(Abcam, ab181975, 1:200),孵育液与封闭液相同。随后,脑膜全片或脑组织切片在 PBS 和 0.05% Tween-20 中室温洗涤三次,每次 10 分钟,再以相同封闭液加入驴源 Alexa Fluor 405、488 或 594 标记的抗大鼠、山羊或兔二抗(Thermo Fisher Scientific, 1:1000)室温孵育 2 小时。对于 Tunel 染色,凋亡细胞检测按照凋亡试剂盒(碧云天生物技术, C1090, 中国)说明书操作。最后,全片在 PBS 中室温洗涤三次,每次 10 分钟。 将组织转移至 PBS 中,并用 ProLong Gold 抗褪色试剂(Invitrogen, P36934 )固定在带有盖玻片的载玻片上。制备好的玻片在 4°C 下保存,并在染色后 1 周内使用 Lecia DMI8 共聚焦显微镜和 LAS AF 软件(Leica Microsystems)进行成像。获取的图像使用 Fiji 软件进行定量分析。对于淋巴结,通过创建淋巴结的三维重建,然后使用 Fiji 计算微珠覆盖的体积除以淋巴结的总体积,来评估清除后的 dCLN 中微珠覆盖的体积百分比。每只小鼠的左右 dCLN 体积百分比取平均值。为了评估脑膜全片中淋巴管(mLV)的覆盖率和复杂性,使用共聚焦显微镜获取脑膜全片的图像,并用 Fiji 进行量化。整个脑膜全片被追踪并用于量化。Lyve1 的面积覆盖率用于确定淋巴管的覆盖率。在适用的情况下,使用相同的图像评估 Lyve1 − CD31 + 血管的视野覆盖率百分比。 所有用于淋巴管形态学定量的脑膜全标本均采用相同的共聚焦设置和 Fiji 参数进行成像。
Flow Cytometric 流式细胞术
Mice were euthanized by Euthasol injection (i.p.) and transcardially perfused with 20 mL cold 1× PBS for 5 min. Heads were removed and skulls were quickly stripped. Mandibles were removed, as well as all skull material rostral to maxillae. Surgical scissors were used to remove the top of the skull, cutting clockwise, beginning and ending inferior to the right post‐tympanic hook. Meninges (dura mater, arachnoid, and pia mater) were carefully removed from the interior aspect of the skulls and surfaces of the brain with Dumont #5 forceps (Fine Science Tools). Meninges were gently pressed through 70‐µm nylon mesh cell strainers with a sterile plastic plunger (BD Biosciences) to yield a single‐cell suspension. For lymphatic endothelial cell isolation, meninges (along with diaphragm and ear skin) were digested for 30 min in 0.60 U mL−1 of Liberase TM (Roche) and 60 U mL−1 of DNase 1. Cells were then centrifuged at 300 RCF at 4 °C for 10 min, the supernatant was removed and cells were resuspended in ice‐cold FACS buffer (pH 7,4; 0.1 m PBS; 1 mm EDTA; 1% BSA). Cells were stained for extracellular marker with antibodies to anti‐CD45‐PE‐Cyanine5 (1:200, clone 30‐F11, eBiosciences), anti‐CD31‐Alexa Fluor 647 (1:200, clone 390, BD Biosciences), and anti‐LYVE1‐ Alexa Fluor 488 (1:200, clone ALY7, eBioscience). Experiments were performed on meninges from n = 2 mice in the sham group and n = 1 mice in the SAH group. For cell apoptosis analysis, the human lymphatic endothelial were plated in a 6‐well plate with 106 cells. After 24 h, the cells were treated with normal medium or conditioned medium (CM) with or without THBS1 antibody (5 µg mL−1, Genetex, GTX21823) for another 48 h. Cells were collected using trypsin–EDTA and resuspended in annexin buffer and incubated for 15 min with Annexin V (BD Biosciences) or PI (eBioscience) according to the manufacturer's protocol. The apoptotic cells were analyzed using a FACScan flow cytometer (BD Biosciences). FACS analyses were reproduced by three independent experiments. Data processing was done with Excel and statistical analysis was performed using GraphPad Prism.
小鼠通过腹腔注射安乐死药物 Euthasol 实施安乐死,随后用 20 mL 冰冷的 1× PBS 进行 5 分钟的心脏灌注。摘除头部并迅速剥离颅骨。去除下颌骨及上颌骨前所有颅骨组织。使用手术剪沿顺时针方向剪开颅顶,起止于右耳后钩下方。用 Dumont #5 镊子(精细科学工具)小心移除颅骨内侧面及脑表面的脑膜(硬脑膜、蛛网膜和软脑膜)。将脑膜通过 70 微米尼龙细胞筛网,用无菌塑料推杆(BD Biosciences)轻柔压过,获得单细胞悬液。为分离淋巴管内皮细胞,将脑膜(连同膈肌和耳部皮肤)置于含 0.60 U/mL Liberase TM(罗氏)和 60 U/mL DNase 1 的溶液中消化 30 分钟。细胞随后在 4°C、300 RCF 条件下离心 10 分钟,弃上清,用冰预冷的 FACS 缓冲液(pH 7.4;0.1 M PBS;1 mM EDTA;1% BSA)重悬细胞。 细胞使用抗 CD45-PE-Cyanine5(1:200,克隆号 30-F11,eBiosciences)、抗 CD31-Alexa Fluor 647(1:200,克隆号 390,BD Biosciences)和抗 LYVE1-Alexa Fluor 488(1:200,克隆号 ALY7,eBioscience)抗体进行细胞外标记染色。实验在假手术组 2 只小鼠和 SAH 组 1 只小鼠的脑膜组织上进行。对于细胞凋亡分析,人淋巴管内皮细胞以 10^2 个细胞接种于 6 孔板中。24 小时后,细胞分别用正常培养基或含或不含 THBS1 抗体(5 µg/mL,Genetex,GTX21823)的条件培养基处理 48 小时。使用胰蛋白酶-EDTA 收集细胞,重悬于 Annexin 缓冲液中,并按制造商说明书与 Annexin V(BD Biosciences)或 PI(eBioscience)孵育 15 分钟。凋亡细胞通过 FACScan 流式细胞仪(BD Biosciences)分析。FACS 分析由三次独立实验重复验证。数据处理使用 Excel 完成,统计分析采用 GraphPad Prism 软件进行。
Primary Meningeal Lymphatic Endothelial Cells Culture
原代脑膜淋巴管内皮细胞培养
For mLEC culture, meninges were dissected from P0‐P1 C57/BL6 mice. To obtain a suspension of meningeal LECs, skull caps were quickly collected, and meninges (dura mater and arachnoid) were dissected using Dumont no. 5 and no. 7 fine forceps in a complete medium composed of Dulbecco's modified Eagle's medium (DMEM) (Gibco) with 2% FBS, 1% l‐glutamine (Gibco), 1% penicillin/streptomycin (Gibco), 1% sodium pyruvate (Gibco), 1% non‐essential amino‐acids (Gibco), and 1.5% Hepes buffer (Gibco). Meninges were then incubated with 1 mL DMEM with 0.60 U mL−1 of Liberase TM (Roche) and 60 U mL−1 of DNase 1 for 30 min at 37 °C. Cell suspensions were then pooled into a single tube after filtration through a 70‐µm nylon mesh cell strainer. Cells were then centrifuged at 300 RCF at 4 °C for 10 min, the supernatant was removed and cells were resuspended with 1 mL ice‐cold magnetic separation buffer. Added 5 µL rat anti‐lyve1(Santa Cruze, sc‐65647, 1:200) antibody to the cell suspension. Vortexed briefly to mix and incubate at 4 °C for 15 min. Washed cells by adding 10 mL of magnetic separation buffer to the tube and centrifuged at 600 RCF for 5 min at 4 °C. Carefully aspirated the supernatant and resuspend the cells in 1 mL of magnetic separation buffer. Added 100 µL of anti‐rat IgG beads (Miltenyi Biotec, cat.no 130‐048‐502) to the cells, vortexed briefly to mix, and incubated for 15 min at 4 °C. Added 10 mL of magnetic separation buffer to the cells and centrifuged at 4 °C, 600 RCF for 5 min. According to the manufacturer's instructions, MACS sorting was carried out by using the magnetic beads matched with the anti‐rat IgG beads. Lyve1‐positive cells were collected and resuspended in mLEC culture medium, inoculated in a culture dish, and the medium was changed every 2 days.
;; 对于 mLEC 培养,从 P0-P1 C57/BL6 小鼠中解剖出脑膜。为获得脑膜淋巴内皮细胞(LECs)悬液,迅速收集颅盖,并使用 Dumont 5 号和 7 号精细镊子在由 Dulbecco 改良 Eagle 培养基(DMEM)(Gibco)配制的完全培养基中解剖脑膜(硬脑膜和蛛网膜),该培养基含 2%胎牛血清(FBS)、1% L-谷氨酰胺(Gibco)、1%青霉素/链霉素(Gibco)、1%丙酮酸钠(Gibco)、1%非必需氨基酸(Gibco)及 1.5% Hepes 缓冲液(Gibco)。随后,将脑膜与 1 mL 含 0.60 U/mL Liberase TM(Roche)和 60 U/mL DNase 1 的 DMEM 在 37°C 下孵育 30 分钟。细胞悬液经 70 微米尼龙网细胞过滤器过滤后合并至单管中。细胞在 4°C 下以 300 RCF 离心 10 分钟,弃去上清,用 1 mL 冰磁分离缓冲液重悬细胞。向细胞悬液中加入 5 µL 大鼠抗 lyve1 抗体(Santa Cruze, sc-65647, 1:200)。短暂涡旋混匀后于 4°C 孵育 15 分钟。加入 10 mL 磁分离缓冲液洗涤细胞,4°C 下以 600 RCF 离心 5 分钟。 小心吸取上清液,将细胞重悬于 1 mL 磁珠分离缓冲液中。加入 100 µL 抗大鼠 IgG 磁珠(Miltenyi Biotec,货号 130-048-502)至细胞悬液,短暂涡旋混匀后于 4°C 孵育 15 分钟。加入 10 mL 磁珠分离缓冲液,4°C、600 RCF 离心 5 分钟。按照制造商说明,使用与抗大鼠 IgG 磁珠匹配的磁珠进行 MACS 分选。收集 Lyve1 阳性细胞,重悬于 mLEC 培养基中接种至培养皿,每 2 天更换一次培养基。
THP1 Conditioned Medium Preparation
THP1 条件培养基制备
Human monocyte THP1 cells were cultured in RPMI 1640 medium containing 10% FBS (VivaCell, Shanghai, China) and 1% penicillin‐streptomycin. For CM collection, THP1 cells were plated in a 6‐well plate with FreeStyle expression medium (12 338 026; Gibco, Thermo Fisher Scientific, USA) for 24 h. The CM were collected 24 h later, centrifuged at 15 000 × g for 5 min, and filtered through a 0.22‐mm filter. Human lymphatic endothelial cells were exposed to fresh medium mixed with CM at a ratio of 1:1 (v/v) for the following process.
人单核细胞 THP1 培养于含 10%胎牛血清(VivaCell,中国上海)及 1%青霉素-链霉素的 RPMI 1640 培养基中。收集条件培养基时,THP1 细胞接种于 6 孔板内 FreeStyle 表达培养基(12 338 026;Gibco,Thermo Fisher Scientific,美国)中培养 24 小时。24 小时后收集上清,15 000×g 离心 5 分钟,经 0.22 µm 滤膜过滤。人淋巴管内皮细胞后续处理采用新鲜培养基与条件培养基按 1:1(体积比)混合的培养基。
Co‐Immunoprecipitation 共免疫沉淀
106 human lymphatic endothelial cells were lysed using 400 µL of lysis buffer (Tris‐HCL HCl 50 mm, pPH 7.4, Nacl NaCl 150 mm, sodium deoxycholate 0.25%, NP‐40 1%, EDTA 1 mm, PMSF 1 mm, Aprotinin 1 mg mL−1, leupeptin 1 mg mL−1, pepstatin 1 mg mL−1) on ice for 30 min and centrifuged at 16 113.6 rcf for 15 min. The supernatant was incubated with CD47 (Invitrogen, MA5‐11895) or IgG (Abclone, AC005) antibodies overnight and followed by protein A/G plus agarose bead (Santa Cruz Biotech., sc‐2003) for another 4 h. After washing three times with ice‐cold PBS, parvalbumin (1 ug mL−1) was added for 2 h. Agarose beads were washed three times with PBS and followed by SDS‐PAGE and immunoblot analysis with indicated antibodies.
10 6 人淋巴管内皮细胞使用 400 µL 裂解缓冲液(Tris-HCL HCl 50 mM,pH 7.4,NaCl 150 mM,脱氧胆酸钠 0.25%,NP-40 1%,EDTA 1 mM,PMSF 1 mM,抑肽酶 1 mg/mL −1 ,亮抑酶肽 1 mg/mL −1 ,胃酶抑素 1 mg/mL −1 )在冰上裂解 30 分钟,随后以 16,113.6 rcf 离心 15 分钟。上清液与 CD47(Invitrogen,MA5-11895)或 IgG(Abclone,AC005)抗体孵育过夜,再加入蛋白 A/G 琼脂糖珠(Santa Cruz Biotech.,sc-2003)继续孵育 4 小时。用冰预冷的 PBS 洗涤三次后,加入小清蛋白(1 µg/mL −1 )孵育 2 小时。琼脂糖珠经 PBS 洗涤三次后,进行 SDS-PAGE 及指定抗体的免疫印迹分析。
Western Blot Analysis 蛋白质印迹分析
Western blot was used to analyze the lymphoid apoptosis pathway‐associated protein changes in meninges at different time points after SAH/sham surgery. Briefly, after euthanasia and perfusion with cold PBS, the mouse meninges were harvested, and total protein was extracted with RIPA lysis buffer and 1% PMSF. The protein was separated by 10% SDS‐PAGE at 120 V for 90 min, and the separated proteins were transferred to PVDF membranes. Then, the PVDF membranes were blocked and incubated with primary antibodies (rabbit anti‐STAT3 (1:1000, Abclone, cat no. A1192, China); rabbit anti‐ Phospho‐STAT3‐Y705 (1:1000, Abclone, cat no.AP0070, China); rabbit anti‐Bcl2 (1:1000, Abcam, cat no. ab182858); rabbit anti‐Bax, (1:1000, Abcam, cat no. ab182734); and rabbit anti‐β actin pAb (1:1000, diagbio, cat no. db10001, China)) on a shaker overnight at 4 °C. After being sufficiently washed, the membranes were incubated with horseradish peroxidase‐conjugated secondary antibodies (1:5000, beyotime, cat no. A0208) for 1 h at room temperature. Experiments were performed on meninges from n = 3 mice per group. A ChemiDoc imaging system (Bio‐Rad) was used to visualize the membranes, and ImageJ software was used to calculate the grey values of the bands.
采用 Western blot 分析 SAH/假手术后不同时间点脑膜淋巴凋亡通路相关蛋白的变化。简言之,小鼠经安乐死及冷 PBS 灌注后,收集脑膜组织,使用 RIPA 裂解液和 1% PMSF 提取总蛋白。蛋白样品经 10% SDS-PAGE(120 V,90 分钟)分离后,转印至 PVDF 膜。随后,PVDF 膜经封闭处理,与一抗(兔抗 STAT3,1:1000,Abclone,货号 A1192,中国;兔抗磷酸化 STAT3-Y705,1:1000,Abclone,货号 AP0070,中国;兔抗 Bcl2,1:1000,Abcam,货号 ab182858;兔抗 Bax,1:1000,Abcam,货号 ab182734;兔抗β actin 多克隆抗体,1:1000,diagbio,货号 db10001,中国)在 4°C 摇床中孵育过夜。充分洗涤后,膜与辣根过氧化物酶标记的二抗(1:5000,beyotime,货号 A0208)室温孵育 1 小时。每组实验使用 n=3 只小鼠的脑膜样本。 使用 ChemiDoc 成像系统(Bio-Rad)观察膜,并用 ImageJ 软件计算条带的灰度值。
Single‐Cell RNA Sequencing
单细胞 RNA 测序
Freshly prepared meningeal cell suspensions were performed immediately according to the manufacturer's protocol of 10× Chromium 3' v3 kit (10× Genomics, Pleasanton, CA). Libraries were sequenced on the Illumina NovaSeq 6000 platform at Novogene, Beijing, China. On average, each sample generated about 709 m reads (SD = 121 m).
新鲜制备的脑膜细胞悬液立即按照 10× Chromium 3' v3 试剂盒(10× Genomics,普莱森顿,加利福尼亚州)制造商的协议进行处理。文库在中国北京诺禾致源的 Illumina NovaSeq 6000 平台上进行测序。平均每个样本产生约 7.09 亿条读长(标准差=1.21 亿)。
Read Alignment, Quality Control, and Clustering Analysis
读段比对、质量控制与聚类分析
The raw gene expression matrix was generated via CellRanger. Doublets were removed through Scrublet.[
56
] Then filtered counts matrix was converted to Seurat object using the Seurat R package (version 4.0.1). The cells with gene numbers >7000, or <200, or >20% mitochondrial counts were filtered. Each sample data was normalized via LogNormalize method and the top 2000 highly variable genes were selected for subsequent dimensional reduction. The batch effect across samples was removed via the integration method implemented in the Seurat package. PCA (n = 30) and the UMAP were used to reduce the dimensionality and visualize the cell distribution. Cluster‐specific genes were identified using the Wilcox rank‐sum test implemented in the FindAllMarkers function (fold change ≥ 2 and adjusted p ≤0.05). Cell clusters were annotated to known cell lineages using well‐recognized marker genes.
原始基因表达矩阵通过 CellRanger 生成。使用 Scrublet 去除双联体细胞。 [
56
] 随后,过滤后的计数矩阵利用 Seurat R 包(版本 4.0.1)转换为 Seurat 对象。筛选掉基因数>7000 或<200,或线粒体计数占比>20%的细胞。各样本数据通过 LogNormalize 方法进行标准化,并选取前 2000 个高变基因用于后续降维分析。样本间的批次效应通过 Seurat 包中的整合方法予以消除。采用 PCA(主成分分析,n=30)和 UMAP 进行降维并可视化细胞分布。通过 FindAllMarkers 函数中的 Wilcox 秩和检验(倍数变化≥2 且校正 p 值≤0.05)鉴定簇特异性基因。利用公认的标志基因将细胞簇注释至已知细胞谱系。
Cell–Cell Communication Analysis
细胞间通讯分析
To analyze cell–cell interactions in the microenvironment, CellChat was used to infer ligand–receptor pairs between mLECs and other cell types.[
57
] In brief, normalized expression values of each sample were fed into CellChat, and identifyOverExpressedGenes, identifyOverExpressedInteractions, as well as projectData functions were used to conduct preprocessing. Core functions “computeCommunProb,” “computeCommunProbPathway,” and “aggregateNet” were applied to infer cell‐cell communication networks. Significant interactions were defined with p ≤ 0.05. Finally, “netAnalysis_signalingRole_heatmap” function was used to determine the senders and receivers in the network.
为分析微环境中的细胞间相互作用,采用 CellChat 推断 mLECs 与其他细胞类型间的配体-受体对。 [
57
] 简言之,将各样本的标准化表达值输入 CellChat,并运用 identifyOverExpressedGenes、identifyOverExpressedInteractions 及 projectData 功能进行预处理。核心功能“computeCommunProb”“computeCommunProbPathway”和“aggregateNet”用于推断细胞间通讯网络。显著性互作定义为 p≤0.05。最后通过“netAnalysis_signalingRole_heatmap”功能确定网络中的信号发送与接收细胞。
Spatial Transcriptomics Sequencing and Data Processing
空间转录组测序与数据处理
Used OCT‐embedded frozen meninge for ST detection. The capture of gene expression information for ST slides was performed by the Visium Spatial platform of 10× Genomics through the use of spatially barcoded mRNA‐binding oligonucleotides in the default protocol. Raw sequencing reads of spatial transcriptomics were quality checked and mapped to the pre‐built mm10 reference genome by Space Ranger v1.1.
使用 OCT 包埋的冷冻脑膜组织进行空间转录组检测。Visium Spatial 平台(10× Genomics)通过默认流程中的空间条形码 mRNA 结合寡核苷酸捕获 ST 玻片的基因表达信息。空间转录组原始测序读数经质量检查后,由 Space Ranger v1.1 比对至预构建的 mm10 参考基因组。
Filtering, Normalization, and Deconvolution of Visium Data
Visium 数据的过滤、标准化与解卷积
The gene‐spot matrices generated after ST data processing were analyzed with the Seurat package (version 4.0.1) in R. Spots were filtered for gene counts <500, mitochondrial counts >25%, or red cell counts >20%. Normalization across spots was performed with the SCTransform function. Dimensionality reduction and clustering were performed with PCA with the top 30 PCs. Spatial feature expression plots were generated with the SpatialFeaturePlot function in Seurat (version 4.0.1). RCTD was used to deconvolute each spot to predict the underlying composition of cell types.[
25
]
ST 数据处理后生成的基因点矩阵使用 R 中的 Seurat 包(版本 4.0.1)进行分析。过滤掉基因计数<500、线粒体计数>25%或红细胞计数>20%的点位。通过 SCTransform 函数进行跨点位标准化。采用前 30 个主成分的 PCA 进行降维和聚类分析。空间特征表达图由 Seurat(版本 4.0.1)中的 SpatialFeaturePlot 函数生成。利用 RCTD 解卷积每个点位以预测潜在的细胞类型组成。 [
25
]
Cell Trajectory Analysis
细胞轨迹分析
The Monocle2 package was used for pseudotime trajectory construction.[
58
] Briefly, the differentialGeneTest function was used to identify DEGs across cell types. The top 1000 genes based on q value were selected as ordering genes. DDRTree method was applied to reduce dimension. orderCells function was used to order cells with the sham group as root.
使用 Monocle2 包构建拟时序轨迹。 [
58
] 简要而言,通过 differentialGeneTest 函数鉴定跨细胞类型的差异表达基因(DEGs)。基于 q 值选取前 1000 个基因作为排序基因。应用 DDRTree 方法进行降维。以假手术组为根,使用 orderCells 函数对细胞进行排序。
Gene Set Enrichment Analysis
基因集富集分析
The marker genes for each group of the mLEC cluster were identified by FindMarkers from Seurat with a threshold of min.pct = 0.1 and logFC.threshold = 0 and then sorted according to avg_log2FC. The GSEA software (v4.1.0) was used to identify the Kyoto Encyclopedia of Genes and Genomes pathways.
通过 Seurat 中的 FindMarkers 功能,以 min.pct = 0.1 和 logFC.threshold = 0 为阈值,识别了 mLEC 簇各组的标志基因,并依据 avg_log2FC 进行排序。随后使用 GSEA 软件(v4.1.0)鉴定了京都基因与基因组百科全书通路。
Statistical Analysis 统计分析
Statistical analysis was performed using SPSS analysis tools (IBM Corp.) or Prism 9 software program (GraphPad Software). All data are presented as the mean ± SEM. To assess the statistical significance between the two groups, a two‐tailed unpaired Student's t‐test (for parametric analysis) or Mann–Whitney U test (for nonparametric analysis) was performed. One‐way analysis of variance (ANOVA) followed by Tukey's or Dunnett's multiple comparisons test were used to detect the differences in the results between groups. The correlation between the expression profiles of CSF biomarkers and the prognosis of SAH patients was analyzed using Spearman's rank test. p values less than 0.05 were considered to indicate statistical significance. No sample outliers were excluded. Individual in vitro experiments were performed at least three times with similar results.
统计分析采用 SPSS 分析工具(IBM Corp.)或 Prism 9 软件(GraphPad Software)进行。所有数据均以均值±标准误表示。为评估两组间的统计学显著性,采用双尾非配对 Student's t 检验(参数分析)或 Mann–Whitney U 检验(非参数分析)。组间结果差异通过单因素方差分析(ANOVA)结合 Tukey 或 Dunnett 多重比较检验进行检测。CSF 生物标志物表达谱与 SAH 患者预后的相关性采用 Spearman 秩检验分析。p 值小于 0.05 视为具有统计学意义。未排除任何样本异常值。体外实验均独立重复至少三次,结果具有一致性。
Ethics Statement 伦理声明
All studies concerning human information and tissues were approved by the Ethics Committee of the Second Affiliated Hospital of Zhejiang University School of Medicine (No.2020‐892) and is registered as an observational study in ClinicalTrail.gov (No. NCT04938414). All of the patients signed informed consent. All animal procedures were approved by the second affiliated hospital of Zhejiang university – Animal Ethics Committee (Approval no. AIRB‐2022‐0698) and were performed according to the Guiding Principles for the Care and Use of Laboratory Animals Approved by Animal Regulations of National Science and Technology Committee of China.
所有涉及人类信息和组织的研究均获得浙江大学医学院附属第二医院伦理委员会批准(编号:2020‐892),并在 ClinicalTrail.gov 注册为观察性研究(编号: NCT04938414 )。所有患者均签署知情同意书。所有动物实验程序经浙江大学附属第二医院动物伦理委员会批准(批准号:AIRB‐2022‐0698),并遵循中国国家科学技术委员会颁布的《实验动物管理条例》中关于实验动物护理与使用的指导原则执行。
Conflict of Interest 利益冲突声明
The authors declare no conflict of interest.
作者声明无利益冲突。
Author Contributions 作者贡献
X.W., A.Z., Q.Y., and Z.W. contributed equally to this work. X.W., A.Z., JM.Z., S.C., and K.W. designed this study. J.L., S.X., JY.Z., Y.Q., J.Z., Y.F., and Y.L. supervised this study. Q.Y., JH.Z., P.X., and J.W. coordinated the sample collection. X.W., Q.Y., H.L., JH.Z., and Y.L. performed all experiments. A.Z., Q.Y., and K.W. contributed to bioinformatics and statistical analysis. S.X. and Z.W. provided technical support for scRNA‐Seq and ST data analysis. X.W., A.Z., Q.Y., and JM.Z. wrote the original manuscript. Y.F., H.L., Y.Q., J.L., K.W., and JW.Z. reviewed and polished the manuscript.
X.W.、A.Z.、Q.Y.和 Z.W.对本工作贡献均等。X.W.、A.Z.、JM.Z.、S.C.和 K.W.设计了本研究。J.L.、S.X.、JY.Z.、Y.Q.、J.Z.、Y.F.和 Y.L.监督了本研究。Q.Y.、JH.Z.、P.X.和 J.W.协调了样本收集。X.W.、Q.Y.、H.L.、JH.Z.和 Y.L.进行了所有实验。A.Z.、Q.Y.和 K.W.参与了生物信息学与统计分析。S.X.和 Z.W.为单细胞 RNA 测序及空间转录组数据分析提供了技术支持。X.W.、A.Z.、Q.Y.和 JM.Z.撰写了初稿。Y.F.、H.L.、Y.Q.、J.L.、K.W.和 JW.Z.审阅并润色了稿件。
Supporting information 支持信息
Supporting Information 支持信息
Acknowledgements 致谢
The authors wish to thank all the patients enrolled in this study. The authors acknowledge that some of the schematics were created using BioRender (https://biorender.com/). This study was supported by grants from the National Natural Science Foundation of China (81870916, 82071287, 82271301) to J.Z., National Natural Science Foundation of China (81971107) to S.C., National Natural Science Foundation of China (82002634) to K.W., National Natural Science Foundation of China (82201430) to Y.F., and National Natural Science Foundation of China (82201512) to S.X.
作者谨向参与本研究的所有患者表示感谢。作者声明部分示意图使用 BioRender( https://biorender.com/ )制作。本研究得到了国家自然科学基金(81870916、82071287、82271301)对 J.Z.的资助,国家自然科学基金(81971107)对 S.C.的资助,国家自然科学基金(82002634)对 K.W.的资助,国家自然科学基金(82201430)对 Y.F.的资助,以及国家自然科学基金(82201512)对 S.X.的资助。
Wang X., Zhang A., Yu Q., Wang Z., Wang J., Xu P., Liu Y., Lu J., Zheng J., Li H., Qi Y., Zhang J., Fang Y., Xu S., Zhou J., Wang K., Chen S., Zhang J., Single‐Cell RNA Sequencing and Spatial Transcriptomics Reveal Pathogenesis of Meningeal Lymphatic Dysfunction after Experimental Subarachnoid Hemorrhage. Adv. Sci.
2023, 10, 2301428. 10.1002/advs.202301428
王 X., 张 A., 于 Q., 王 Z., 王 J., 徐 P., 刘 Y., 卢 J., 郑 J., 李 H., 齐 Y., 张 J., 方 Y., 徐 S., 周 J., 王 K., 陈 S., 张 J., 单细胞 RNA 测序与空间转录组学揭示实验性蛛网膜下腔出血后脑膜淋巴功能障碍的发病机制。《先进科学》2023 年第 10 期,2301428 号。DOI: 10.1002/advs.202301428
Contributor Information 贡献者信息
Kaikai Wang, Email: wangkaikai@zju.edu.cn.
王凯凯,电子邮箱:wangkaikai@zju.edu.cn。
Sheng Chen, Email: saintchan@zju.edu.cn.
陈晟,电子邮箱:saintchan@zju.edu.cn。
Jianmin Zhang, Email: zjm135@zju.edu.cn.
张建民,Email: zjm135@zju.edu.cn。
Data Availability Statement
数据可用性声明
The data that support the findings of this study are available from the corresponding author upon reasonable request.
支持本研究结果的数据可根据合理请求从通讯作者处获取。
References 参考文献
-
1.
Feng D., Zhou J., Liu H., Wu X., Li F., Zhao J., Zhang Y., Wang L., Chao M., Wang Q., Qin H., Ge S., Liu Q., Zhang J., Qu Y., Sci. Adv.
2022, 8, eabq2423. [DOI] [PMC free article] [PubMed] [Google Scholar]
1. 冯东, 周杰, 刘辉, 吴晓, 李飞, 赵静, 张洋, 王力, 晁明, 王强, 秦浩, 葛松, 刘青, 张军, 曲阳, 《科学进展》2022 年 8 卷, eabq2423. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
2.
Athiraman U., Zipfel G. J., Int. J. Mol. Sci.
2021, 22, 6550. [DOI] [PMC free article] [PubMed] [Google Scholar]
2. Athiraman U., Zipfel G. J., 《国际分子科学杂志》2021 年,第 22 卷,6550 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
3.a) Fang Y., Wang X., Lu J., Shi H., Huang L., Shao A., Zhang A., Liu Y., Ren R., Lenahan C., Tang J., Zhang J., Zhang J. H., Chen S., EBioMedicine
2022, 76, 103843; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Chen J., Wang L., Xu H., Xing L., Zhuang Z., Zheng Y., Li X., Wang C., Chen S., Guo Z., Liang Q., Wang Y., Nat. Commun.
2020, 11, 3159. [DOI] [PMC free article] [PubMed] [Google Scholar]
3.a) 方勇,王晓,卢杰,史浩,黄磊,邵安,张安,刘洋,任荣,Lenahan C.,唐杰,张杰,张建宏,陈松,《EBioMedicine》2022 年,第 76 卷,103843 页; [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]; b) 陈杰,王磊,徐浩,邢磊,庄志,郑阳,李翔,王超,陈松,郭志,梁强,王勇,《自然通讯》2020 年,第 11 卷,3159 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
4.a) Alitalo K., Nat. Med.
2011, 17, 1371; [DOI] [PubMed] [Google Scholar]; b) Aspelund A., Robciuc M. R., Karaman S., Makinen T., Alitalo K., Circ. Res.
2016, 118, 515; [DOI] [PubMed] [Google Scholar]; c) Wiig H., Swartz M. A., Physiol. Rev.
2012, 92, 1005. [DOI] [PubMed] [Google Scholar]
4.a) Alitalo K., 《自然医学》2011 年,第 17 卷,1371 页; [ DOI ] [ PubMed ] [ Google Scholar ]; b) Aspelund A., Robciuc M. R., Karaman S., Makinen T., Alitalo K., 《循环研究》2016 年,第 118 卷,515 页; [ DOI ] [ PubMed ] [ Google Scholar ]; c) Wiig H., Swartz M. A., 《生理学评论》2012 年,第 92 卷,1005 页。 [ DOI ] [ PubMed ] [ Google Scholar ] -
5.a) Ahn J. H., Cho H., Kim J. H., Kim S. H., Ham J. S., Park I., Suh S. H., Hong S. P., Song J. H., Hong Y. K., Jeong Y., Park S. H., Koh G. Y., Nature
2019, 572, 62; [DOI] [PubMed] [Google Scholar]; b) Louveau A., Smirnov I., Keyes T. J., Eccles J. D., Rouhani S. J., Peske J. D., Derecki N. C., Castle D., Mandell J. W., Lee K. S., Harris T. H., Kipnis J., Nature
2015, 523, 337; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Aspelund A., Antila S., Proulx S. T., Karlsen T. V., Karaman S., Detmar M., Wiig H., Alitalo K., J. Exp. Med.
2015, 212, 991. [DOI] [PMC free article] [PubMed] [Google Scholar]
5.a) Ahn J. H., Cho H., Kim J. H., Kim S. H., Ham J. S., Park I., Suh S. H., Hong S. P., Song J. H., Hong Y. K., Jeong Y., Park S. H., Koh G. Y., 《自然》2019 年,572 卷,62 页;[ DOI ] [ PubMed ] [ Google Scholar ];b) Louveau A., Smirnov I., Keyes T. J., Eccles J. D., Rouhani S. J., Peske J. D., Derecki N. C., Castle D., Mandell J. W., Lee K. S., Harris T. H., Kipnis J., 《自然》2015 年,523 卷,337 页;[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ];c) Aspelund A., Antila S., Proulx S. T., Karlsen T. V., Karaman S., Detmar M., Wiig H., Alitalo K., 《实验医学杂志》2015 年,212 卷,991 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
6.a) Mesquita S. D., Louveau A., Vaccari A., Smirnov I., Cornelison R. C., Kingsmore K. M., Contarino C., Onengut‐Gumuscu S., Farber E., Raper D., Viar K. E., Powell R. D., Baker W., Dabhi N., Bai R., Cao R., Hu S., Rich S. S., Munson J. M., Lopes M. B., Overall C. C., Acton S. T., Kipnis J., Nature
2018, 560, 185; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Ma Q., Ineichen B. V., Detmar M., Proulx S. T., Nat. Commun.
2017, 8, 1434. [DOI] [PMC free article] [PubMed] [Google Scholar]
6.a) Mesquita S. D., Louveau A., Vaccari A., Smirnov I., Cornelison R. C., Kingsmore K. M., Contarino C., Onengut‐Gumuscu S., Farber E., Raper D., Viar K. E., Powell R. D., Baker W., Dabhi N., Bai R., Cao R., Hu S., Rich S. S., Munson J. M., Lopes M. B., Overall C. C., Acton S. T., Kipnis J., 《自然》2018 年,560 卷,185 页;[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ];b) Ma Q., Ineichen B. V., Detmar M., Proulx S. T., 《自然通讯》2017 年,8 卷,1434 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
7.
Oliver G., Kipnis J., Randolph G. J., Harvey N. L., Cell
2020, 182, 270. [DOI] [PMC free article] [PubMed] [Google Scholar]
7. Oliver G., Kipnis J., Randolph G. J., Harvey N. L., 《细胞》2020 年,182 卷,270 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
8.a) Chen H., Ye F., Guo G., Cell Mol. Immunol.
2019, 16, 242; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Xin Z., Lin M., Hao Z., Chen D., Chen Y., Chen X., Xu X., Li J., Wu D., Chai Y., Wu P., Nat. Commun.
2022, 13, 5463. [DOI] [PMC free article] [PubMed] [Google Scholar]
8.a) 陈 H., 叶 F., 郭 G., 《细胞与分子免疫学》2019, 16, 242; [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]; b) 辛 Z., 林 M., 郝 Z., 陈 D., 陈 Y., 陈 X., 徐 X., 李 J., 吴 D., 柴 Y., 吴 P., 《自然通讯》2022, 13, 5463. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
9.a) Lein E., Borm L. E., Linnarsson S., Science
2017, 358, 64; [DOI] [PubMed] [Google Scholar]; b) Nerurkar S. N., Goh D., Cheung C. C. L., Nga P. Q. Y., Lim J. C. T., Yeong J. P. S., Cancers
2020, 12, 2572. [DOI] [PMC free article] [PubMed] [Google Scholar]
9.a) Lein E., Borm L. E., Linnarsson S., 《科学》2017 年,358 卷,64 页;[ DOI ] [ PubMed ] [ Google Scholar ];b) Nerurkar S. N., Goh D., Cheung C. C. L., Nga P. Q. Y., Lim J. C. T., Yeong J. P. S., 《癌症》2020 年,12 卷,2572 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
10.
Zhu Q., Shah S., Dries R., Cai L., Yuan G. C., Nat. Biotechnol.
2018, 36, 1183. [DOI] [PMC free article] [PubMed] [Google Scholar]
10. Zhu Q., Shah S., Dries R., Cai L., Yuan G. C., 《自然·生物技术》2018 年,36 卷,1183 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
11.a) Moncada R., Barkley D., Wagner F., Chiodin M., Devlin J. C., Baron M., Hajdu C. H., Simeone D. M., Yanai I., Nat. Biotechnol.
2020, 38, 333; [DOI] [PubMed] [Google Scholar]; b) Liu Y. M., Ge J. Y., Chen Y. F., Liu T., Chen L., Liu C. C., Ma D., Chen Y. Y., Cai Y. W., Xu Y. Y., Shao Z. M., Yu K. D., Adv. Sci. (Weinheim, Ger.)
2023, 10, 2205395. [Google Scholar]
11.a) Moncada R., Barkley D., Wagner F., Chiodin M., Devlin J. C., Baron M., Hajdu C. H., Simeone D. M., Yanai I., 《自然·生物技术》2020 年,38 卷,333 页;[ DOI ] [ PubMed ] [ Google Scholar ];b) Liu Y. M., Ge J. Y., Chen Y. F., Liu T., Chen L., Liu C. C., Ma D., Chen Y. Y., Cai Y. W., Xu Y. Y., Shao Z. M., Yu K. D., 《先进科学》(德国魏因海姆)2023 年,10 卷,2205395 页。[ Google Scholar ] -
12.
Stuart T., Butler A., Hoffman P., Hafemeister C., Papalexi E., Mauck W. M. 3rd, Hao Y., Stoeckius M., Smibert P., Satija R., Cell
2019, 177, 1888. [DOI] [PMC free article] [PubMed] [Google Scholar]
12. Stuart T., Butler A., Hoffman P., Hafemeister C., Papalexi E., Mauck W. M. 3rd, Hao Y., Stoeckius M., Smibert P., Satija R., 《细胞》2019 年,177 卷,1888 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
13.
Martin J. C., Chang C., Boschetti G., Ungaro R., Giri M., Grout J. A., Gettler K., Chuang L. S., Nayar S., Greenstein A. J., Dubinsky M., Walker L., Leader A., Fine J. S., Whitehurst C. E., Mbow M. L., Kugathasan S., Denson L. A., Hyams J. S., Friedman J. R., Desai P. T., Ko H. M., Laface I., Akturk G., Schadt E. E., Salmon H., Gnjatic S., Rahman A. H., Merad M., Cho J. H., et al., Cell
2019, 178, 1493. [DOI] [PMC free article] [PubMed] [Google Scholar]
13. Martin J. C., Chang C., Boschetti G., Ungaro R., Giri M., Grout J. A., Gettler K., Chuang L. S., Nayar S., Greenstein A. J., Dubinsky M., Walker L., Leader A., Fine J. S., Whitehurst C. E., Mbow M. L., Kugathasan S., Denson L. A., Hyams J. S., Friedman J. R., Desai P. T., Ko H. M., Laface I., Akturk G., Schadt E. E., Salmon H., Gnjatic S., Rahman A. H., Merad M., Cho J. H., 等,《细胞》2019 年,第 178 卷,第 1493 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
14.
Qi J., Crinier A., Escalière B., Ye Y., Wang Z., Zhang T., Batista L., Liu H., Hong L., Wu N., Zhang M., Chen L., Liu Y., Shen L., Narni‐Mancinelli E., Vivier E., Su B., Cell Rep. Med.
2021, 2, 100353. [DOI] [PMC free article] [PubMed] [Google Scholar]
14. Qi J., Crinier A., Escalière B., Ye Y., Wang Z., Zhang T., Batista L., Liu H., Hong L., Wu N., Zhang M., Chen L., Liu Y., Shen L., Narni‐Mancinelli E., Vivier E., Su B., 《细胞报告·医学》2021 年,第 2 卷,第 100353 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
15.
Wong H. Y., Sheng Q., Hesterberg A. B., Croessmann S., Rios B. L., Giri K., Jackson J., Miranda A. X., Watkins E., Schaffer K. R., Donahue M., Winkler E., Penson D. F., Smith J. A., Herrell S. D., Luckenbaugh A. N., Barocas D. A., Kim Y. J., Graves D., Giannico G. A., Rathmell J. C., Park B. H., Gordetsky J. B., Hurley P. J., Nat. Commun.
2022, 13, 6036. [DOI] [PMC free article] [PubMed] [Google Scholar]
15. Wong H. Y., Sheng Q., Hesterberg A. B., Croessmann S., Rios B. L., Giri K., Jackson J., Miranda A. X., Watkins E., Schaffer K. R., Donahue M., Winkler E., Penson D. F., Smith J. A., Herrell S. D., Luckenbaugh A. N., Barocas D. A., Kim Y. J., Graves D., Giannico G. A., Rathmell J. C., Park B. H., Gordetsky J. B., Hurley P. J., 《自然·通讯》2022 年,第 13 卷,第 6036 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
16.
Xiao Y., Cong M., Li J., He D., Wu Q., Tian P., Wang Y., Yang S., Liang C., Liang Y., Wen J., Liu Y., Luo W., Lv X., He Y., Cheng D. D., Zhou T., Zhao W., Zhang P., Zhang X., Xiao Y., Qian Y., Wang H., Gao Q., Yang Q. C., Yang Q., Hu G., Cancer Cell
2021, 39, 423. [DOI] [PubMed] [Google Scholar]
16. 肖勇,丛明,李杰,何东,吴强,田鹏,王洋,杨松,梁超,梁燕,文静,刘洋,罗伟,吕鑫,何毅,程大东,周涛,赵伟,张鹏,张晓,肖勇,钱阳,王浩,高强,杨启成,杨强,胡刚,《癌细胞》2021 年,第 39 卷,第 423 页。[ DOI ] [ PubMed ] [ Google Scholar ] -
17.
Bajpai G., Bredemeyer A., Li W., Zaitsev K., Koenig A. L., Lokshina I., Mohan J., Ivey B., Hsiao H. M., Weinheimer C., Kovacs A., Epelman S., Artyomov M., Kreisel D., Lavine K. J., Circ. Res.
2019, 124, 263. [DOI] [PMC free article] [PubMed] [Google Scholar]
17. 巴杰派,布雷德梅耶,李伟,扎伊采夫,柯尼格,洛克希娜,莫汉,艾维,萧海明,温海默,科瓦奇,埃佩尔曼,阿尔乔莫夫,克赖塞尔,拉维恩,《循环研究》2019 年,第 124 卷,第 263 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
18.
Zhang L., Yu X., Zheng L., Zhang Y., Li Y., Fang Q., Gao R., Kang B., Zhang Q., Huang J. Y., Konno H., Guo X., Ye Y., Gao S., Wang S., Hu X., Ren X., Shen Z., Ouyang W., Zhang Z., Nature
2018, 564, 268. [DOI] [PubMed] [Google Scholar]
18. 张磊,余鑫,郑磊,张洋,李毅,方强,高然,康斌,张琪,黄俊阳,今野博,郭晓,叶阳,高松,王松,胡鑫,任鑫,沈哲,欧阳伟,张哲,《自然》2018 年,第 564 卷,第 268 页。[ DOI ] [ PubMed ] [ Google Scholar ] -
19.
Sawai C. M., Sisirak V., Ghosh H. S., Hou E. Z., Ceribelli M., Staudt L. M., Reizis B., J. Exp. Med.
2013, 210, 2151. [DOI] [PMC free article] [PubMed] [Google Scholar]
19. 泽维·萨维,西斯拉克,戈什,侯恩泽,塞里巴利,斯托特,雷齐斯,《实验医学杂志》2013 年,第 210 卷,第 2151 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
20.
Ben‐Baruch Morgenstern N., Ballaban A. Y., Wen T., Shoda T., Caldwell J. M., Kliewer K., Felton J. M., Abonia J. P., Mukkada V. A., Putnam P. E., Bolton S. M., Dwyer D. F., Barrett N. A., Rothenberg M. E., J. Allergy Clin. Immunol.
2022, 149, 2062. [DOI] [PMC free article] [PubMed] [Google Scholar]
20. Ben‐Baruch Morgenstern N., Ballaban A. Y., Wen T., Shoda T., Caldwell J. M., Kliewer K., Felton J. M., Abonia J. P., Mukkada V. A., Putnam P. E., Bolton S. M., Dwyer D. F., Barrett N. A., Rothenberg M. E., 《过敏与临床免疫学杂志》2022 年,第 149 卷,第 2062 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
21.
Brown H. M., Anastasi M. R., Frank L. A., Kind K. L., Richani D., Robker R. L., Russell D. L., Gilchrist R. B., Thompson J. G., Biol. Reprod.
2015, 92, 26. [DOI] [PubMed] [Google Scholar]
21. Brown H. M., Anastasi M. R., Frank L. A., Kind K. L., Richani D., Robker R. L., Russell D. L., Gilchrist R. B., Thompson J. G., 《生殖生物学》2015 年,第 92 卷,第 26 页。 [ DOI ] [ PubMed ] [ Google Scholar ] -
22.
Morimoto J., Matsumoto M., Miyazawa R., Oya T., Tsuneyama K., Matsumoto M., Immunohorizons
2022, 6, 202. [DOI] [PubMed] [Google Scholar]
22. Morimoto J., Matsumoto M., Miyazawa R., Oya T., Tsuneyama K., Matsumoto M., 《免疫视野》2022 年,第 6 卷,第 202 页。 [ DOI ] [ PubMed ] [ Google Scholar ] - 23. Jung S., Lee M., Kim D. Y., Son C., Ahn B. H., Heo G., Park J., Kim M., Park H. E., Koo D. J., Park J. H., Lee J. W., Choe H. K., Kim S. Y., Neuron 2022, 110, 266. [DOI] [PubMed] [Google Scholar]
-
24.
Smillie C. S., Biton M., Ordovas‐Montanes J., Sullivan K. M., Burgin G., Graham D. B., Herbst R. H., Rogel N., Slyper M., Waldman J., Sud M., Andrews E., Velonias G., Haber A. L., Jagadeesh K., Vickovic S., Yao J., Stevens C., Dionne D., Nguyen L. T., Villani A. C., Hofree M., Creasey E. A., Huang H., Rozenblatt‐Rosen O., Garber J. J., Khalili H., Desch A. N., Daly M. J., Ananthakrishnan A. N., et al., Cell
2019, 178, 714. [DOI] [PMC free article] [PubMed] [Google Scholar]
24. Smillie C. S., Biton M., Ordovas‐Montanes J., Sullivan K. M., Burgin G., Graham D. B., Herbst R. H., Rogel N., Slyper M., Waldman J., Sud M., Andrews E., Velonias G., Haber A. L., Jagadeesh K., Vickovic S., Yao J., Stevens C., Dionne D., Nguyen L. T., Villani A. C., Hofree M., Creasey E. A., Huang H., Rozenblatt‐Rosen O., Garber J. J., Khalili H., Desch A. N., Daly M. J., Ananthakrishnan A. N., 等, 《细胞》2019 年, 178 卷, 714 页. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] - 25. Cable D. M., Murray E., Zou L. S., Goeva A., Macosko E. Z., Chen F., Irizarry R. A., Nat. Biotechnol. 2022, 40, 517. [DOI] [PMC free article] [PubMed] [Google Scholar]
-
26.a) Zeyu Z., Yuanjian F., Cameron L., Sheng C., Exp. Neurol.
2021, 336, 113535; [DOI] [PubMed] [Google Scholar]; b) Jin J., Duan J., Du L., Xing W., Peng X., Zhao Q., Front. Immunol.
2022, 13, 1027756; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Geraghty J. R., Lung T. J., Hirsch Y., Katz E. A., Cheng T., Saini N. S., Pandey D. K., Testai F. D., Neurosurgery
2021, 89, 1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
26.a) 泽宇 Z., 元健 F., 卡梅伦 L., 盛 C., 《实验神经学》2021 年, 336 卷, 113535 页; [ DOI ] [ PubMed ] [ Google Scholar ]; b) 金 J., 段 J., 杜 L., 邢 W., 彭 X., 赵 Q., 《免疫学前沿》2022 年, 13 卷, 1027756 页; [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]; c) 杰拉蒂 J. R., 朗 T. J., 赫希 Y., 卡茨 E. A., 程 T., 赛尼 N. S., 潘迪 D. K., 泰斯塔伊 F. D., 《神经外科》2021 年, 89 卷, 1071 页. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
27.
Pham D., Tan X., Xu J., Grice L. F., Lam P. Y., Raghubar A., Vukovic J., Ruitenberg M. J., Nguyen Q., bioRxiv
2020, 2020.05.31.125658. [Google Scholar]
27. 范 D., 谭 X., 徐 J., 格莱斯 L. F., 林 P. Y., 拉古巴尔 A., 武科维奇 J., 鲁滕伯格 M. J., 阮 Q., 《生物预印本》2020 年, 2020.05.31.125658. [ Google Scholar ] -
28.
Plana E., Oto J., Medina P., Herranz R., Fernández‐Pardo Á., Requejo L., Miralles M., IUBMB Life
2022, 74, 982. [DOI] [PubMed] [Google Scholar]
28. 普拉纳 E., 奥托 J., 梅迪纳 P., 赫兰兹 R., 费尔南德斯‐帕尔多 Á., 雷克霍 L., 米拉莱斯 M., 《国际生物化学与分子生物学联盟生命》2022 年, 74 卷, 982 页. [ DOI ] [ PubMed ] [ Google Scholar ] -
29.a) Isenberg J. S., Martin‐Manso G., Maxhimer J. B., Roberts D. D., Nat. Rev. Cancer
2009, 9, 182; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Kaur S., Martin‐Manso G., Pendrak M. L., Garfield S. H., Isenberg J. S., Roberts D. D., J. Biol. Chem.
2010, 285, 38923. [DOI] [PMC free article] [PubMed] [Google Scholar]
29.a) 伊森伯格 J. S., 马丁‐曼索 G., 马克西默 J. B., 罗伯茨 D. D., 《自然评论癌症》2009 年, 9 卷, 182 页; [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]; b) 考尔 S., 马丁‐曼索 G., 彭德拉克 M. L., 加菲尔德 S. H., 伊森伯格 J. S., 罗伯茨 D. D., 《生物化学杂志》2010 年, 285 卷, 38923 页. [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] - 30. Subramanian A., Tamayo P., Mootha V. K., Mukherjee S., Ebert B. L., Gillette M. A., Paulovich A., Pomeroy S. L., Golub T. R., Lander E. S., Mesirov J. P., Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 15545. [DOI] [PMC free article] [PubMed] [Google Scholar]
-
31.
Liu K., Ren T., Huang Y., Sun K., Bao X., Wang S., Zheng B., Guo W., Cell Death Dis.
2017, 8, e3015. [DOI] [PMC free article] [PubMed] [Google Scholar]
31. 刘凯,任涛,黄毅,孙科,鲍晓,王松,郑波,郭伟,《细胞死亡与疾病》2017 年,第 8 卷,e3015。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
32.
Huang Y. C., Chang C. Y., Wu Y. Y., Wu K. L., Tsai Y. M., Lee H. C., Tsai E. M., Hsu Y. L., Cancers
2022, 15, 176.36612175 [Google Scholar]
32. 黄永成,张春阳,吴洋洋,吴克亮,蔡一鸣,李汉成,蔡恩明,徐玉兰,《癌症》2022 年,第 15 卷,176.36612175 [ Google Scholar ] -
33.
Bulters D., Gaastra B., Zolnourian A., Alexander S., Ren D., Blackburn S. L., Borsody M., Doré S., Galea J., Iihara K., Nyquist P., Galea I., Nat. Rev. Neurol.
2018, 14, 416. [DOI] [PubMed] [Google Scholar]
33. 布尔特斯 D.,加斯特拉 B.,佐尔努里安 A.,亚历山大 S.,任 D.,布莱克本 S.L.,博索迪 M.,多雷 S.,加利亚 J.,饭原 K.,尼奎斯特 P.,加利亚 I.,《自然评论·神经病学》2018 年,第 14 卷,416 页。[ DOI ] [ PubMed ] [ Google Scholar ] -
34.
Zoerle T., Lombardo A., Colombo A., Longhi L., Zanier E. R., Rampini P., Stocchetti N., Crit. Care Med.
2015, 43, 168. [DOI] [PubMed] [Google Scholar]
34. 佐尔勒 T.,隆巴尔多 A.,科伦坡 A.,隆吉 L.,扎尼尔 E.R.,兰皮尼 P.,斯托凯蒂 N.,《重症监护医学》2015 年,第 43 卷,168 页。[ DOI ] [ PubMed ] [ Google Scholar ] -
35.
Cahill J., Calvert J. W., Zhang J. H., J. Cereb. Blood Flow Metab.
2006, 26, 1341. [DOI] [PubMed] [Google Scholar]
35. Cahill J., Calvert J. W., Zhang J. H., 《脑血流与代谢杂志》2006 年,第 26 卷,第 1341 页。 [ DOI ] [ PubMed ] [ Google Scholar ] -
36.a) Nagel R. L., Gibson Q. H., J. Biol. Chem.
1971, 246, 69; [PubMed] [Google Scholar]; b) Zhao X., Sun G., Ting S. M., Song S., Zhang J., Edwards N. J., Aronowski J., J. Neurochem.
2015, 133, 144. [DOI] [PMC free article] [PubMed] [Google Scholar]
36.a) Nagel R. L., Gibson Q. H., 《生物化学杂志》1971 年,第 246 卷,第 69 页; [ PubMed ] [ Google Scholar ]; b) Zhao X., Sun G., Ting S. M., Song S., Zhang J., Edwards N. J., Aronowski J., 《神经化学杂志》2015 年,第 133 卷,第 144 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
37.a) Galea I., Bechmann I., Perry V. H., Trends Immunol.
2007, 28, 12; [DOI] [PubMed] [Google Scholar]; b) Carson M. J., Doose J. M., Melchior B., Schmid C. D., Ploix C. C., Immunol. Rev.
2006, 213, 48. [DOI] [PMC free article] [PubMed] [Google Scholar]
37.a) Galea I., Bechmann I., Perry V. H., 《免疫学趋势》2007 年,第 28 卷,第 12 页; [ DOI ] [ PubMed ] [ Google Scholar ]; b) Carson M. J., Doose J. M., Melchior B., Schmid C. D., Ploix C. C., 《免疫学评论》2006 年,第 213 卷,第 48 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
38.a) Bower N. I., Hogan B. M., J. Mol. Med.
2018, 96, 383; [DOI] [PubMed] [Google Scholar]; b) Antila S., Karaman S., Nurmi H., Airavaara M., Voutilainen M. H., Mathivet T., Chilov D., Li Z., Koppinen T., Park J. H., Fang S., Aspelund A., Saarma M., Eichmann A., Thomas J. L., Alitalo K., J. Exp. Med.
2017, 214, 3645. [DOI] [PMC free article] [PubMed] [Google Scholar]
38.a) Bower N. I., Hogan B. M., 《分子医学杂志》2018 年,第 96 卷,第 383 页; [ DOI ] [ PubMed ] [ Google Scholar ]; b) Antila S., Karaman S., Nurmi H., Airavaara M., Voutilainen M. H., Mathivet T., Chilov D., Li Z., Koppinen T., Park J. H., Fang S., Aspelund A., Saarma M., Eichmann A., Thomas J. L., Alitalo K., 《实验医学杂志》2017 年,第 214 卷,第 3645 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
39.
Coles J. A., Myburgh E., Brewer J. M., McMenamin P. G., Prog. Neurobiol.
2017, 156, 107. [DOI] [PubMed] [Google Scholar]
39. 科尔斯 J. A., 米堡 E., 布鲁尔 J. M., 麦克梅纳明 P. G., 《神经生物学进展》2017 年, 156 卷, 107 页。 [ DOI ] [ PubMed ] [ Google Scholar ] -
40.a) Bolte A. C., Dutta A. B., Hurt M. E., Smirnov I., Kovacs M. A., McKee C. A., Ennerfelt H. E., Shapiro D., Nguyen B. H., Frost E. L., Lammert C. R., Kipnis J., Lukens J. R., Nat. Commun.
2020, 11, 4524; [DOI] [PMC free article] [PubMed] [Google Scholar]; b) Liu X., Gao C., Yuan J., Xiang T., Gong Z., Luo H., Jiang W., Song Y., Huang J., Quan W., Wang D., Tian Y., Ge X., Lei P., Zhang J., Jiang R., Acta Neuropathol. Commun.
2020, 8, 16. [DOI] [PMC free article] [PubMed] [Google Scholar]
40.a) 博尔特 A. C., 杜塔 A. B., 赫特 M. E., 斯米尔诺夫 I., 科瓦奇 M. A., 麦基 C. A., 恩纳菲特 H. E., 夏皮罗 D., 阮 B. H., 弗罗斯特 E. L., 拉默特 C. R., 基普尼斯 J., 卢肯斯 J. R., 《自然通讯》2020 年, 11 卷, 4524 页; [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]; b) 刘 X., 高 C., 袁 J., 向 T., 龚 Z., 罗 H., 姜 W., 宋 Y., 黄 J., 全 W., 王 D., 田 Y., 葛 X., 雷 P., 张 J., 姜 R., 《神经病理学通讯》2020 年, 8 卷, 16 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
41.
Liu Q., Hou C., Zhang H., Fu C., Wang W., Wang B., Li J., Zhao Y., Yang X., Brain Res.
2021, 1769, 147584. [DOI] [PubMed] [Google Scholar]
41. 刘强,侯晨,张华,付超,王伟,王斌,李杰,赵阳,杨旭,《脑研究》2021 年,1769 卷,147584 页。[ DOI ] [ PubMed ] [ Google Scholar ] -
42.a) de Lima K. A., Rustenhoven J., Kipnis J., Annu. Rev. Immunol.
2020, 38, 597; [DOI] [PubMed] [Google Scholar]; b) Rustenhoven J., Drieu A., Mamuladze T., de Lima K. A., Dykstra T., Wall M., Papadopoulos Z., Kanamori M., Salvador A. F., Baker W., Lemieux M., Mesquita S. Da, Cugurra A., Fitzpatrick J., Sviben S., Kossina R., Bayguinov P., Townsend R. R., Zhang Q., Erdmann‐Gilmore P., Smirnov I., Lopes M. B., Herz J., Kipnis J., Cell
2021, 184, 1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
42.a) 德利马·K·A,拉斯滕霍文·J,基普尼斯·J,《免疫学年评》2020 年,38 卷,597 页;[ DOI ] [ PubMed ] [ Google Scholar ];b) 拉斯滕霍文·J,德里乌·A,马穆拉德兹·T,德利马·K·A,戴克斯特拉·T,沃尔·M,帕帕多普洛斯·Z,卡纳莫里·M,萨尔瓦多·A·F,贝克·W,勒米厄·M,梅斯基塔·S·达,库古拉·A,菲茨帕特里克·J,斯维本·S,科西纳·R,贝吉诺夫·P,汤森德·R·R,张强,埃尔德曼-吉尔莫·P,斯米尔诺夫·I,洛佩斯·M·B,赫兹·J,基普尼斯·J,《细胞》2021 年,184 卷,1000 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
43.
Wolf F. W., Eddy R. L., Shows T. B., Dixit V. M., Genomics
1990, 6, 685. [DOI] [PubMed] [Google Scholar]
43. 沃尔夫·F·W,埃迪·R·L,肖斯·T·B,迪克西特·V·M,《基因组学》1990 年,6 卷,685 页。[ DOI ] [ PubMed ] [ Google Scholar ] -
44.
Huang T., Sun L., Yuan X., Qiu H., OncoTargets Ther.
2017, 8, 84546. [DOI] [PMC free article] [PubMed] [Google Scholar]
44. 黄涛,孙亮,袁翔,邱海,《肿瘤靶向治疗》2017 年,8 卷,84546 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
45.
Jeanne A., Schneider C., Martiny L., Dedieu S., Front. Pharmacol.
2015, 6, 252. [DOI] [PMC free article] [PubMed] [Google Scholar]
45. Jeanne A., Schneider C., Martiny L., Dedieu S., 《前沿药理学》2015 年第 6 卷,第 252 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
46.
Kamijo H., Miyagaki T., Takahashi‐Shishido N., Nakajima R., Oka T., Suga H., Sugaya M., Sato S., Leukemia
2020, 34, 845. [DOI] [PubMed] [Google Scholar]
46. Kamijo H., Miyagaki T., Takahashi‐Shishido N., Nakajima R., Oka T., Suga H., Sugaya M., Sato S., 《白血病》2020 年第 34 卷,第 845 页。 [ DOI ] [ PubMed ] [ Google Scholar ] -
47.
Horiguchi H., Yamagata S., Rong Qian Z., Kagawa S., Sakashita N., J. Med. Invest.
2013, 60, 91. [DOI] [PubMed] [Google Scholar]
47. Horiguchi H., Yamagata S., Rong Qian Z., Kagawa S., Sakashita N., 《医学研究杂志》2013 年第 60 卷,第 91 页。 [ DOI ] [ PubMed ] [ Google Scholar ] -
48.
Pal S. K., Nguyen C. T., Morita K. I., Miki Y., Kayamori K., Yamaguchi A., Sakamoto K., J. Oral Pathol. Med.
2016, 45, 730. [DOI] [PubMed] [Google Scholar]
48. Pal S. K., Nguyen C. T., Morita K. I., Miki Y., Kayamori K., Yamaguchi A., Sakamoto K., 《口腔病理学与医学杂志》2016 年第 45 卷,第 730 页。 [ DOI ] [ PubMed ] [ Google Scholar ] - 49. Donato R., Cannon B. R., Sorci G., Riuzzi F., Hsu K., Weber D. J., Geczy C. L., Curr. Mol. Med. 2013, 13, 24. [PMC free article] [PubMed] [Google Scholar]
-
50.a) Graczyk A., Leśniak W., Int. J. Biochem. Cell Biol.
2014, 57, 135; [DOI] [PubMed] [Google Scholar]; b) Donato R., Sorci G., Giambanco I., Cell. Mol. Life Sci.
2017, 74, 2749; [DOI] [PMC free article] [PubMed] [Google Scholar]; c) Gross S. R., Sin C. G., Barraclough R., Rudland P. S., Cell. Mol. Life Sci.
2014, 71, 1551. [DOI] [PMC free article] [PubMed] [Google Scholar]
50.a) Graczyk A., Leśniak W., 《国际生物化学与细胞生物学杂志》2014 年,57 卷,135 页;[ DOI ] [ PubMed ] [ Google Scholar ];b) Donato R., Sorci G., Giambanco I., 《细胞与分子生命科学》2017 年,74 卷,2749 页;[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ];c) Gross S. R., Sin C. G., Barraclough R., Rudland P. S., 《细胞与分子生命科学》2014 年,71 卷,1551 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
51.
Fang Y., Shao A., Wang X., Lu J., Wu H., Ren R., Huang Y., Lenahan C., Xu J., Chen S., Zhang J., Eur. Radiol.
2021, 31, 1290. [DOI] [PubMed] [Google Scholar]
51. Fang Y., Shao A., Wang X., Lu J., Wu H., Ren R., Huang Y., Lenahan C., Xu J., Chen S., Zhang J., 《欧洲放射学》2021 年,31 卷,1290 页。[ DOI ] [ PubMed ] [ Google Scholar ] -
52.
Li J., Miao B., Wang S., Dong W., Xu H., Si C., Wang W., Duan S., Lou J., Bao Z., Zeng H., Yang Z., Cheng W., Zhao F., Zeng J., Liu X. S., Wu R., Shen Y., Chen Z., Chen S., Wang M., Brief Bioinform.
2022, 23, bbac261. [DOI] [PubMed] [Google Scholar]
52. 李杰,缪波,王松,董伟,徐浩,司超,王伟,段帅,娄静,鲍志,曾华,杨志,程伟,赵飞,曾杰,刘晓生,吴瑞,沈阳,陈志,陈松,王明,《生物信息学简报》2022 年,第 23 卷,bbac261。[ DOI ] [ PubMed ] [ Google Scholar ] -
53.
Schallner N., Pandit R., LeBlanc R. 3rd, Thomas A. J., Ogilvy C. S., Zuckerbraun B. S., Gallo D., Otterbein L. E., Hanafy K. A., J. Clin. Invest.
2015, 125, 2609. [DOI] [PMC free article] [PubMed] [Google Scholar]
53. Schallner N., Pandit R., LeBlanc R. 3rd, Thomas A. J., Ogilvy C. S., Zuckerbraun B. S., Gallo D., Otterbein L. E., Hanafy K. A., 《临床研究杂志》2015 年,125 卷,2609 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
54.
Fang Y., Shi H., Ren R., Huang L., Okada T., Lenahan C., Gamdzyk M., Travis Z. D., Lu Q., Tang L., Huang Y., Zhou K., Tang J., Zhang J., Zhang J. H., Neurotherapeutics
2020, 17, 1954. [DOI] [PMC free article] [PubMed] [Google Scholar]
54. Fang Y., Shi H., Ren R., Huang L., Okada T., Lenahan C., Gamdzyk M., Travis Z. D., Lu Q., Tang L., Huang Y., Zhou K., Tang J., Zhang J., Zhang J. H., 《神经治疗学》2020 年,17 卷,1954 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
55.
Susaki E. A., Tainaka K., Perrin D., Yukinaga H., Kuno A., Ueda H. R., Nat. Protoc.
2015, 10, 1709. [DOI] [PubMed] [Google Scholar]
55. 须崎英明,田中健一,佩兰·D.,雪永悠,久野亚美,上田宏,《自然·实验手册》2015 年,第 10 卷,第 1709 页。[ DOI ] [ PubMed ] [ Google Scholar ] -
56.
Wolock S. L., Lopez R., Klein A. M., Cell Syst.
2019, 8, 281. [DOI] [PMC free article] [PubMed] [Google Scholar]
56. 沃洛克·S. L.,洛佩兹·R.,克莱因·A. M.,《细胞系统》2019 年,第 8 卷,第 281 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
57.
Jin S., Guerrero‐Juarez C. F., Zhang L., Chang I., Ramos R., Kuan C. H., Myung P., Plikus M. V., Nie Q., Nat. Commun.
2021, 12, 1088. [DOI] [PMC free article] [PubMed] [Google Scholar]
57. 金松,格雷罗-胡亚雷斯·C. F.,张亮,常毅,拉莫斯·R.,关志宏,朴明,普利库斯·M. V.,聂青,《自然·通讯》2021 年,第 12 卷,第 1088 页。[ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ] -
58.
Trapnell C., Cacchiarelli D., Grimsby J., Pokharel P., Li S., Morse M., Lennon N. J., Livak K. J., Mikkelsen T. S., Rinn J. L., Nat. Biotechnol.
2014, 32, 381. [DOI] [PMC free article] [PubMed] [Google Scholar]
58. Trapnell C., Cacchiarelli D., Grimsby J., Pokharel P., Li S., Morse M., Lennon N. J., Livak K. J., Mikkelsen T. S., Rinn J. L., 《自然生物技术》2014 年,第 32 卷,第 381 页。 [ DOI ] [ PMC free article ] [ PubMed ] [ Google Scholar ]
Associated Data 相关数据
This section collects any data citations, data availability statements, or supplementary materials included in this article.
本部分收集了本文中包含的任何数据引用、数据可用性声明或补充材料。
Supplementary Materials 补充材料
Supporting Information 支持信息
Data Availability Statement
数据可用性声明
The data that support the findings of this study are available from the corresponding author upon reasonable request.
支持本研究结果的数据可根据合理要求从通讯作者处获取。